(Ckickするとその年度の卒業生の氏名と研究タイトルが表示されます。更にClickすると詳細な研究内容が表示されます。)
2008年度
真正粘菌変形体の成長に伴って形成する 輸送管ネットワーク成長解析 伊藤 昌明
さまざまな問題を理解するための枠組みとして,「複雑ネットワーク」が注目されている.一見複雑に見える現象も,ネットワークの視点から単純な法則を導き出すことができる.人間関係における“六次の隔たり”や,経済活動などにおける“80対20の法則”などがその例である.そしてこの「複雑ネットワーク」は生命における複雑な仕組みや現象を理解する手助けもしてくれるのである.私の研究では真正粘菌変形体をいう生物の生命活動の仕組みを,複雑ネットワーク理論を用いて調べている.
真正粘菌変形体とは,多核の単細胞生物である.単細胞でありながら,その大きさは数μm~数10cmにまで成長し,通常の細胞が10μm程度であることと比べると,非常に大きな細胞であることが分かる.そして,この生物は成長に伴って管の構造を形成し,その管の中では原形質が流動している.すなわち,管は物質を輸送する経路になっている.この非常に大きな組織を維持し,生命活動を行っていくという点で,輸送管がとても重要な役割を果たしていると言えるであろう.
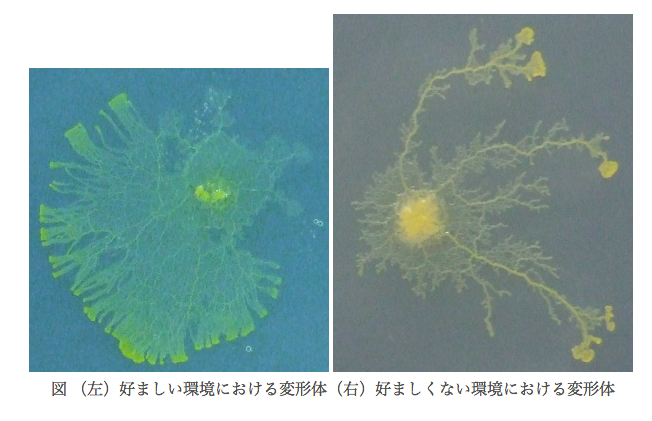
また,変形体は周囲の環境によってその形状を変化させる.それに伴って輸送管のネットワーク構造も変化する.今までの研究で分かっている範囲では,変形体にとって好ましい環境では輸送管ネットワークは格子に近い構造をとり,好ましくない環境では木に近い構造をとる.ここで不思議なのは,このような環境に対する反応を示すにも関わらず,変形体は神経系を持たないことである.
このような変形体の現象を理解し,再構築するために,私は輸送管に注目し,そのネットワークを調べるという手段をとり,現在研究を進めている.
[1] アルバート=ラズロ・バラバシ(青木薫 訳), 新ネットワーク思考~世界の仕組みを 読み解く~, NHK出版 (2002)
[2] 増田直紀, 今野紀雄, 複雑ネットワークの科学, 産業図書 (2005)
生体肝組織を模擬した細胞培養デバイスの開発 海津 新
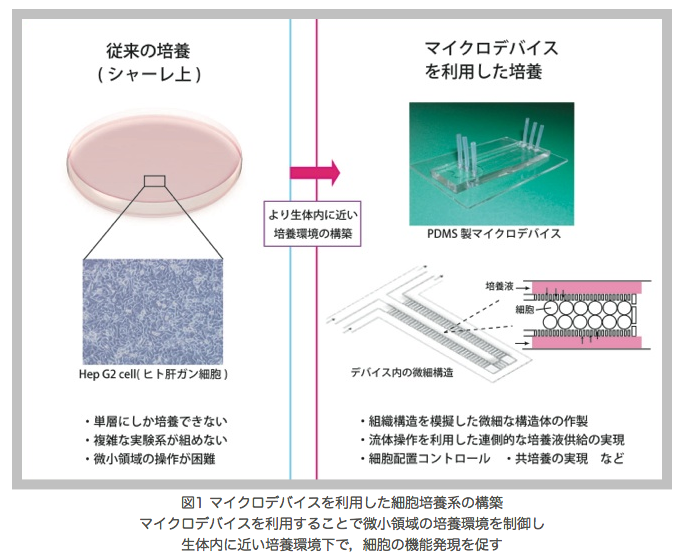
生体外でおこなう細胞培養系において、生体内に近い正常な機能を保持した細胞の獲得は重要な課題である。私たちはマイクロデバイスを利用した細胞培養系を構築することによってこの課題に取り組んでいる。マイクロデバイスはマイクロメートルオーダーの微細な構造を含んだ実験容器である。これまでに、高密度培養のためのマイクロ構造物の設計や栄養分の連続的な供給・老廃物の除去をおこなう灌流システムの開発、他種細胞の共培養法の開発など、従来の培養皿を用いた培養では実現できなかった生体内を模倣する培養環境の構築が研究されている。本研究では、生体内の組織構造を模擬したマイクロ構造の設計や細胞配置、血管系を模倣した栄養分の供給法などを考案し、細胞が持つ特徴的な機能や形態の発現を促すマイクロデバイスの開発を目指している。現在は、肝臓における特徴的な組織構造を模擬し、肝実質細胞間にできる毛細胆管の形成や胆汁の分泌などを促すことを目標にしている。 (共同研究先:東京大学生産技術研究所 マイクロメカトロニクス国際研究センター 藤井(輝)研究室)
2009年度
真正粘菌変形体における パターン形成と環境適応性に関する研究 横谷 佳世子
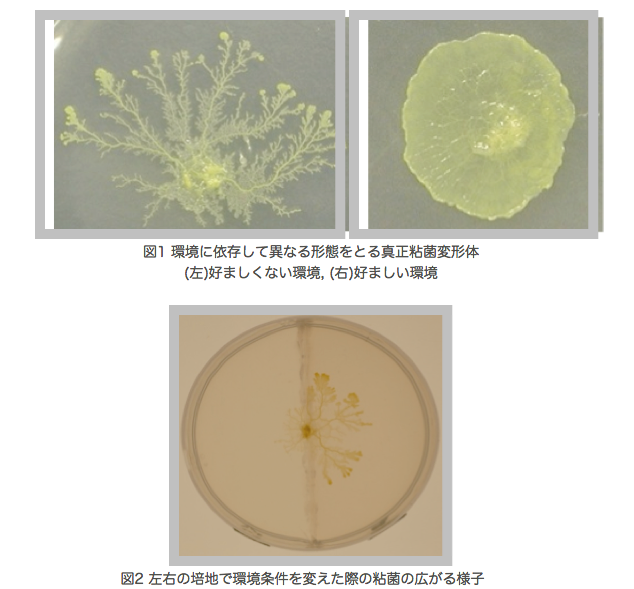
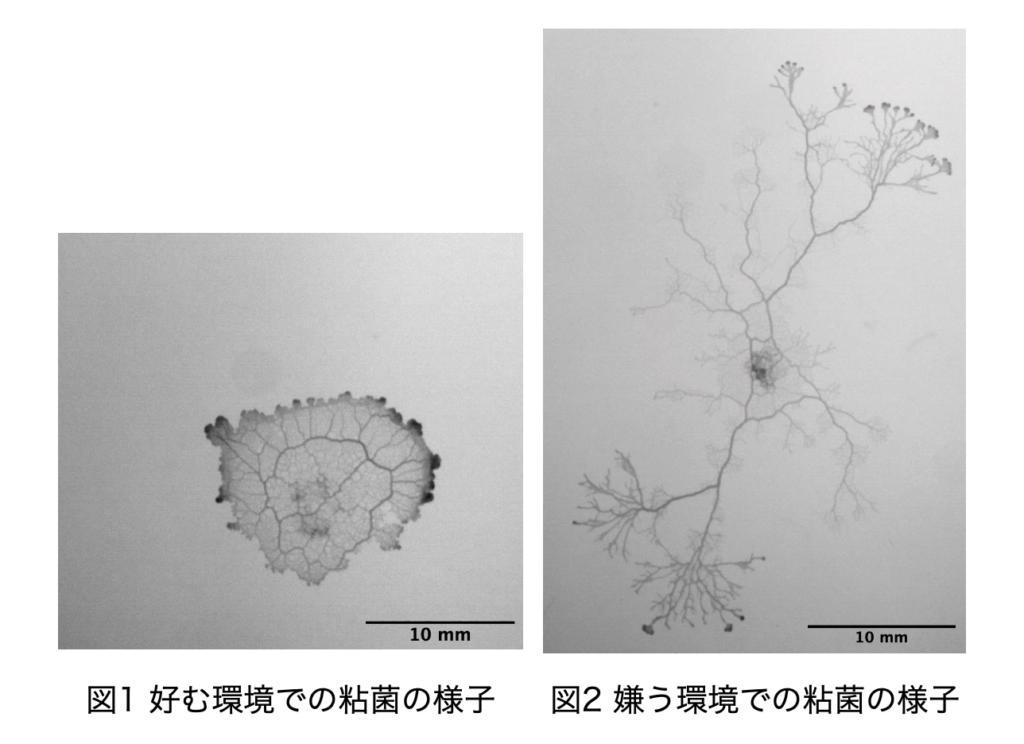
生物は自身が置かれた環境に適応することができる。それは、脳による情報処理の結果であったり、細胞間の複雑な相互作用の結果によるものと考えられるが、本研究で扱っている真正粘菌変形体は神経系も分化構造も持たないにも関わらず、高等生物と同じように周囲の環境に適応できる。真正粘菌変形体とはアメーバ状の多核単細胞生物であり、周囲の環境に応じて自身の形態を変化させることが報告されていて、図1のように好環境時はシート状に、嫌環境時は樹状に広がりながら成長する[1]。本研究では、寒天培地の濃度、培地内の化学物質濃度を組み合わせて変化させ、粘菌にとっての好環境・嫌環境の条件下で形成するパターンの時系列変化を観察している。さらに、特徴的な形状パターンを定量的に解析するために、粘菌の広がる面積や周囲長、複雑度、フラクタル次元などの計測を行っている。最終的には、得られた形状特徴量に基づいて粘菌の環境適応性を定量的に評価するための評価関数を定義し、検証実験によって妥当性の高い評価式を目指す。
[1] Environment-dependent morphology in plasmodium of true slime mold Physarum polycephalum and a network growth model Atsuko Takamatsu , Eri Takaba and Ginjiro Takizawa(2008
真正粘菌変形体におけるアロメトリックスケール則 平井 友

生物のサイズ(M)と基礎代謝量(B)の間にはB=aM^b(a:種などに固有の定、b:ベキ係数)というアロメトリック・スケール則が観察されている。さらにこの関係におけるベキ係数bについて、ネットワークの幾何学的次元数を考慮した理論的予測がなされている[1]。
本研究で用いる真正粘菌変形体は、管状ネットワークを持つ多核単細胞生物であり、細胞融合することによって数10μm 〜 数mまでと非常に幅広いサイズ範囲をもつ性質がある。そして、自然状態では2次元平面状にネットワークを形成するが、環境制御下では1次元形状へとネットワーク構造を変化させることのできる大変興味深い生物である。そのため、真正粘菌変形体は単一種においてアロメトリック・スケール則の検証を行うことができ、さらにその関係におけるベキ係数が幾何学的次元数によるという理論的予測について検証を行うことのできる大変有用な生物であるといえる。そこで、本研究では2次元形状と1次元形状のネットワーク構造をもつ真正粘菌変形体のサイズ(M)と代謝量(B)の間にそれぞれアロメトリック・スケール則が観察されるのか、またベキ係数は幾何学的次元数によるのかを検証していく。さらに、形状変化が与える代謝量への影響について明らかにしていきたい。
[1] Geoffrey B. West, A General Model for the Origin of Allometric Scaling Laws in Biology, SCIENCE VOL. 276(1997)
現場型微生物遺伝子解析装置による定量PCRの実現 平賀 雅隆
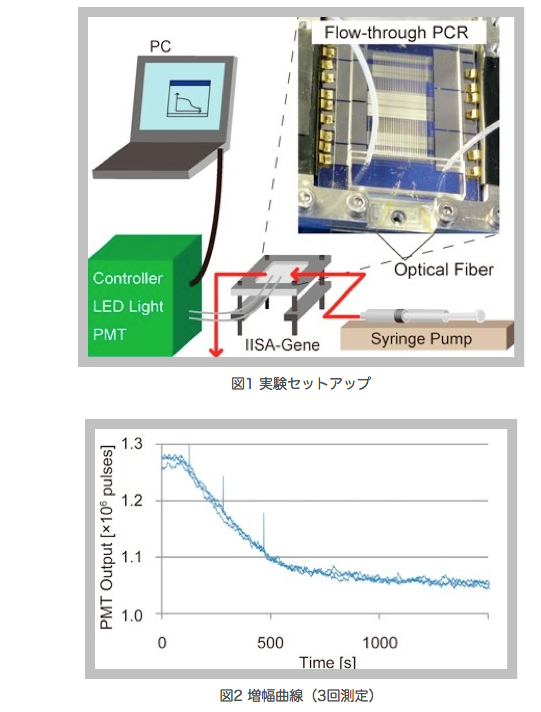
海洋環境の微生物調査を目的として現場型微生物遺伝子解析装置”IISA-Gene” が開発されている.この装置は深海における動作試験で遺伝子の増幅に成功しているが[1],より正確な遺伝子解析を実現するためには,増幅された遺伝子断片の特異性に関する評価が必要である.また,遺伝子の定量を実現できれば,微生物相に関してより詳細な情報を得ることができる.
そこで本研究では,TaqManプローブを用いて,連続流動(Flow-through)方式による定量PCRの実現を目指す.Flow-through PCRの原理は複数の温度帯域を構成し,その上に繰り返し構造を持つ1本の流路を設け,溶液を入口から出口にかけて流すことでPCR温度サイクルを繰り返すものである.溶液出口付近に蛍光検出部を設け,DNA量を蛍光により測定することができる(図1)[2].光検出器(PMT)をアナログ式から高感度・低ノイズのフォトンカウンティング式のものに交換することで,TaqMan プローブの微弱な蛍光変化を計測できるようにした.
実験ではFlow-throug PCRによる増幅が定常状態になったところで,温度設定を変えることでPCRを停止し,そのまま溶液を流し続けることでTaqMan PCRによる遺伝子増幅曲線が得られた(図2).
生物振動子の位相応答曲線の測定とモデル化 松本 秀明
真正粘菌変形体はアメーバのような多核単細胞生物である。
変形体は数10cmもの大きさに成長することもあるが、そのサイズにも関わらず、細胞全体が協同して、変形したり移動したりする。
変形体は、細胞骨格であるアクチン繊維の状態に応じて、1~2分の周期で厚みを振動させている。この厚み振動によって、管状構造を通じた原形質の往復流動が起き、変形体の「部分」どうしは相互作用する。結果として、細胞全体では図2のように様々な厚み振動パターンが計測される。
このような振動パターンを生み出す相互作用を定量的に評価することが、この研究の目的である。
そのため、位相モデルを用いて変形体を結合振動子系として表し[1]、実験的に位相応答曲線[2]を求めている。
最近では、相互作用を評価するための実験手法がいくつも提案されているが、その中の1つである重み付きスパイクトリガー平均法[3]についても、変形体への適用を検討している。
[1] 蔵本由紀編,非線形・非平衡現象の数理1 リズム現象の世界,東京大学出版会, (2005).
[2] C.H.Johnson, Phase Response Curve: What Can They Tell Us about Circadian Clocks?, edited by T.Hiroshige and K.Honma, Hokkaido Univ. Press, Sapporo, 209-246, (2005).
[3] K. Ota, M. Nomura, and T. Aoyagi, A weighted spike-triggered average of a fluctuating stimulus yielding the phase response curve, Phys. Rev. Lett., 024101 (2009).
2010年度
真正粘菌変形体における位相応答曲線の測定 木下 瑛翔
2つの振動する物体(例えば振り子)を考える。今、この2つの振動子がばらばらなタイミングで振動しているとする。ここで、この2つの振動子間に相互作用を与えてあげたとき、その作用によって同位相(つまりタイミングをそろえて)で振動するようになるという現象がおこる。
ところで、この現象を記述しようとしたとき、振動子間に働く相互作用のファクターは無数となることも多い。もしn個のファクターがかかわるとすれば、これはn次元で記述されることになる。しかし、これらを正確に記述するのは現実的に困難なため、「位相縮約」という理論を用いて、一変数φにこの現象を代表させて記述することにする。
本研究では、粘菌断片を振動子として考え、それを2つ結合させた2結合体を2つの相互作用する振動子として考える。粘菌の振動子としての作用を知るために、ここではインパルス刺激を粘菌に与えることで、その反応を確認し、振動子としての粘菌のシステムの性質を理解することが目的である。
しかし、インパルス刺激を加えその反応を見ることは現実的に困難である。そこで本研究ではWSTA(重み付スパイクトリガー法)という方法を用いる。これは、振動子に対して常に刺激を入れ続け、その刺激の形状からインパルス刺激を入れた場合の反応を逆算するという方法である。
粘菌はその培地によって形状を変化させる生き物であるため、培地により振動子の性質が変化しているのではないか、と予想される。本研究では、今後そのことを確認していきたいと思っている。
真正粘菌変形体におけるアメーバ運動の解析 田代 杏子
高等生物は「神経」という非常に複雑なシステムを持っており、その情報処理に基づき様々な行動をしていると考えられる。一方、本研究で扱う真正粘菌変形体は巨大なアメーバ状の単細胞生物である。これは神経系がないにも関わらず、餌を探索しそれを認識するとその部位に集合し、有害な刺激からは逃避するなどの行動をすることができる。単純な構造からどのようにして複雑な行動を生成するのかということを明らかにしていくため、本研究ではアメーバ運動に着目して解析を行う。
ここで、重要となるのが『接触角』というものである。アメーバのような細胞が環境中を這い回るとき、培地に対して接着因子をアンカーとして固定し、前進すると考えられている。アンカーを固定できるかどうかは、細胞と培地の付着力で決まり、それは細胞と培地の間の接触角で見積もることができる。本研究では、様々な環境下で接触角を計測し、粘菌が環境中をどのようにして這い回るのかを明らかにすることを目指す。
真正粘菌変形体の成長に伴って形成する 粘菌ネットワーク成長解析 岡本 力
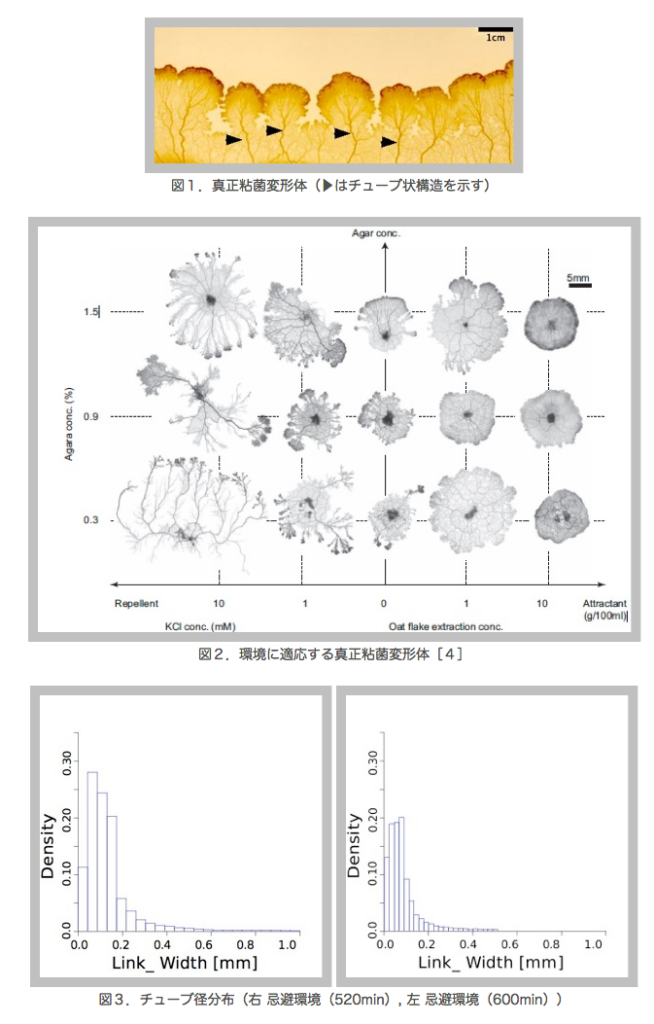
生物は,酸素や栄養分を行き渡らせるために輸送管ネットワークを形成している.動物でいえば,血管ネットワークであり,植物でいえば葉脈や道管である [1].個体群まで拡張すれば,蟻補食時のトレイルパターン [2],鉄道網や道路網 [3] など我々の周りには様々なネットワークが存在している.本研究では,真正粘菌変形体 Physarum polycephalum という巨大なアメーバ様の多核単細胞生物を例に輸送管ネットワークの形態による環境適応機構を探る.
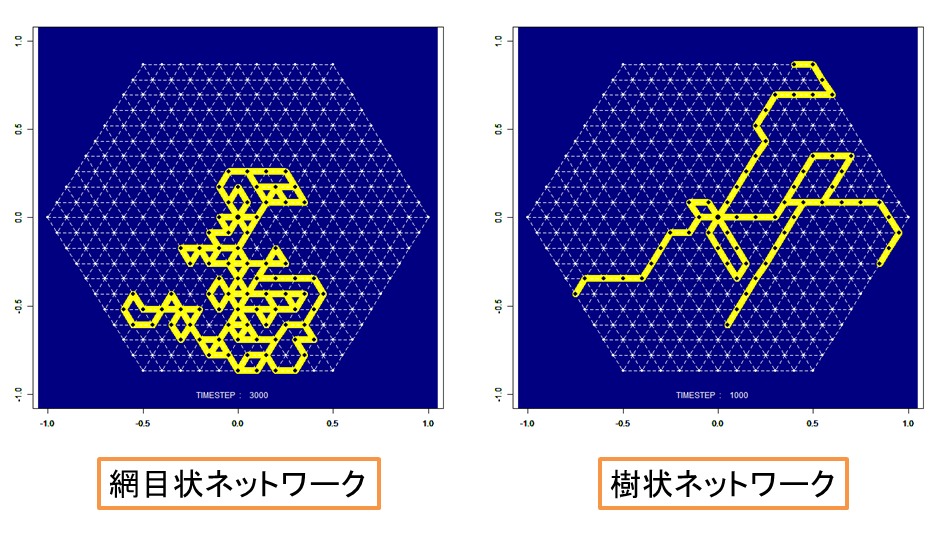
真正粘菌変形体は,多核単細胞生物であり,大きさは数10μmー数10cmまで成長する非常に大きな細胞である.細胞を2つに切り分けても別々の個体として生存でき,逆に,2つの細胞を接触させると1つの細胞に融合して行動ができるのは多核単細胞生物であるからである.また,成長に伴ってチューブ状構造を形成し,その中を原形質が往復流動することにより,細胞全体に栄養分,酸素などを運搬している(図1).また,真正粘菌変形体は神経を持たないが,原形質の往復流動が神経系の代わりに情報伝達を担っているとも考えられる.さらに,真正粘菌は培養する環境に応じて形状が著しく変化する事がわかっている[4](図2).真正粘菌変形体を栄養分が豊富で表面が堅固な培地(誘引環境)で培養した場合には,全体の形状は薄いシート状となり,ネットワークの形態はメッシュ状となる.一方,忌避物質を含ませた培地や表面が柔らかい培地(忌避環境)で培養した場合には,樹状のネットワークが形成される.これらのネットワーク形態は培地条件(栄養分濃度,忌避物質濃度,培地の堅さ)に依存して連続的に変化する.そこで,本研究ではまずこのチューブ状構造変化を定量的に解析を行った.
伊藤らによって,ネットワークのトポロジーのみに着目した研究が行われていた[5].しかし,この研究ではチューブの形状(太さ,長さ)を無視している.ネットワークの物質輸送の効率を考える際,チューブの長さ,太さが重要である.そこで,本研究では各環境下で形成されるネットワーク中でのチューブ径の分布,チューブ径(図3)とチューブ長の分布を定量化した.
今後は,ネットワークが成長する過程で見られる,チューブの生成・消滅のメカニズムを解明を目指す.
[1] Nelson, T. and Dengler N., Leaf cascular pattern formation.The Plant Cell, 9, 1121-1135 (1997).
[2] Burton J. L., and Franks, N. R., The foraging ecology of the army ant Ection rapax An Ergonomic enigrama? Ecol. Entomol. 10, 131-141 (1985)[3] Barth´elemy, M. and Flammini A., Modeling urban street patterns.Phys. Rev. Lett, 100, 138702 (2008).
[4] Takamatsu, A. Takaba, E. Takizawa,G.Environment-dependent morphology in plassmodium of true slime mold Physarum polycephalum and a network growth model.J.Theor. Biol., 256, 29-44 (2009)
[5] 伊藤昌明,高松敦子,生物の輸送管ネットワークの環境適応機構から学ぶ,第51回自動 制御連合講演会,336-340,(2009)
2011年度
真正粘菌変形体の形状の違いによる ミトコンドリアの量の変化 佐々木 崇人
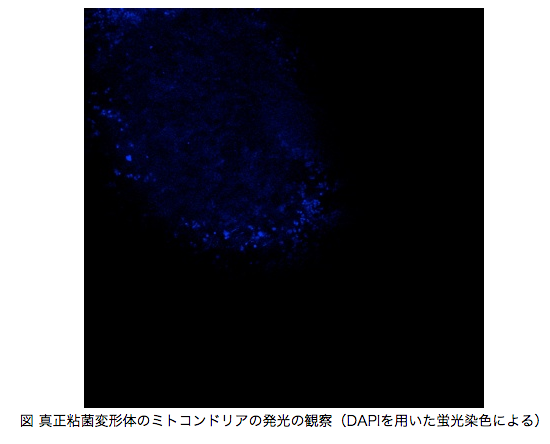
真正粘菌変形体は多核の単細胞生物であり,その大きさは数10μm〜数mまでと非常に幅広いサイズ範囲に成長するが, 切断しても別々の個体として生きていくことが出来る.また,成長に伴って管の構造を形成することで,細胞全体で高度なネットワークを形成する.さらに環境によって形状が異なり,誘引環境ではシート状に,忌避環境では樹状にそれぞれ広がることが知られている.このような形状の違いにより,酸素消費量が異なることが先行研究により分かっている[1].
ここでは,酸素消費量に関わるミトコンドリアに着目し,形状によってミトコンドリアの量を変化させることで,粘菌のような単純な生命体でも環境に応じた効率的な選択を可能にしていると考えた.実験では,ミトコンドリアの量を測定するためにDAPIという蛍光染色で染色し,共焦点レーザー顕微鏡で観察・撮影を行った[4](図).
本研究では,このミトコンドリアの量の観察を進めるとともに,実験から忌避環境における理想的なネットワークを考察し,現実系に落としこむことを目指す.
[1] T.Kuroiwa, S.Kawano, M.Hizume, ”Studies on mitochondrial structure and function in Physarum polycephalum. V. Behaviour of mitochondrial Nucleoids throughout Mitochondrial Division Cycle”, J cell Biol., 72(3), 687-694 (1977)
[2] Y.Moriyama, T.Yamazaki, H.Nomura, N.Sasaki, S.Kawano ”Early zygote- specific nuclease in mitochondria of the true smile mold Physarum polycephalum”, Curr Genet., 48, 334-343 (2005)
[3] Lincoln V. Johnson., Maria L. Walsh., Lan Bo Chen., ”Localization of mitochondria in living cells with rhodamine 123”, Broc. Natl. Acad. sci. USA., 77, 990-994 (1980)
[4] S.Hirose, A.Komamine, ”Observations of Mitochondria and Mitochondrial Nuclei by Double Staining with Rhodamine-123 and DAPI in Synchronous Cultures of Catharanthus roseus”, Bot. Mag. Tokyo., 105, 405-411(1992)
結合振動子系における生命現象のダイナミクスと ネットワーク形状の関係 嶋谷 拓弥
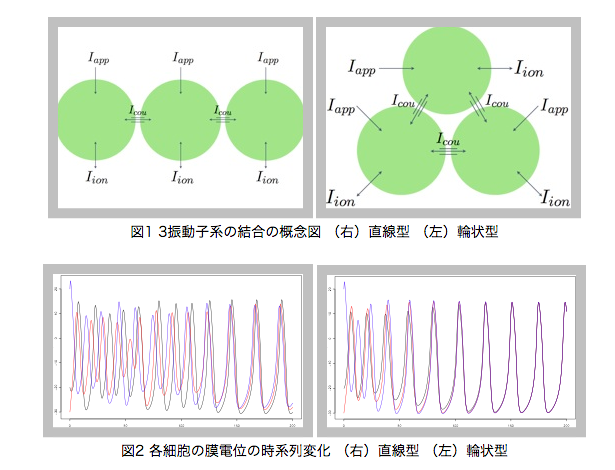
生命現象は複数の反応が様々な形で繋がり合い,反応系全体で複雑なネットワークを構成している.個々の反応では振動現象が見られ,結合が見られる部分では相互作用による干渉が見られるが,全体としては安定な系である.これは反応系の複雑なネットワークがフィードバックシステムを構成し,系全体で安定になるよう制御しているためであると考えられている.現実の反応では生体分子の拡散や各過程の反応時間などのため,系に遅れ時間が発生する.この遅れ時間を考慮したシステムを時間遅れフィードバックシステムといい,複雑な生命現象をより現実系に近い形で表すシステムとして考えられている.
ここではネットワーク形状が系の複雑な反応を生み出す鍵であると考え,システムの形状を変化させることでそのダイナミクスがいかに変化するかに着目した.ギャップ結合という細胞間結合で起こる反応過程を数理的視点から考察した Morris-Lecar モデルに対してシミュレーションを行い,ネットワーク形状ごとに振動現象を観察してその同調現象を比較した.(図1,図2)
本研究では,ネットワーク形状と系の分岐構造の関係の調査を進めるとともに,上述の時間遅れフィードバックの概念を考慮してより現実系に近い生命システムのモデルを構築し,最終的には複雑ネットワークを構成する生命システムのメカニズムや安定性を解明することを目指す.
真正粘菌変形体における形状と酸素消費量の関係 遠藤 辰弥
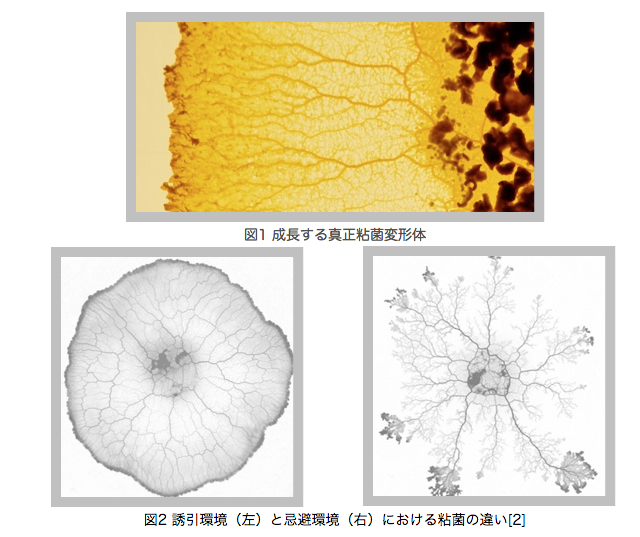
図1に示すように生物の体重(M)と基礎代謝量(B)がある関係に従ってるとことが観察されている.この関係はB=aM^b(a:種などに固有の定数,b:ベキ係数)で表されており,アロメトリック・スケール則と呼ばれている.この関係におけるべき係数bに対し,ネットワークの幾何学的次元数Dを考慮した理論的予測がなされている[1].
真正粘菌変形体(以下「粘菌」)は多核の単細胞生物であり,体内に管を生成して栄養分や酸素などを細胞全体に運搬することで移動・成長をする(図1).さらに硬い培地など粘菌にとって好条件の場合は円形に近い形状となり,一方でやわらかい培地など粘菌にとって悪条件の場合にはひも状に近い形状となる(図2).
先行研究では,五味によって粘菌の酸素消費量とフラクタル次元の間には相関があり,フラクタル次元が小さくなると酸素消費量も小さくなることが示唆された[3].また,平井によって粘菌が円形に近い形状のとき,粘菌の酸素消費量と体重(M)の間に関係があることが明らかになり,理論的な予測として生物を構成する管が細い場合には,「ネットワークの幾何学的次元数Dである生物に対して,基礎代謝量(B)は体重(M)の3D/(2D+3)乗に比例する」というモデルが考案された[4].
本研究では,先行研究の「代謝量(B)が次元数(D)にどのように依存するのか」をさらに追究するとともに,粘菌の形状は代謝量に影響を与えるのかを明らかにすることを目的としている.なおこの研究を行うにあたっては,代謝量の指標として酸素消費量を,次元数の指標としてフラクタル次元をそれぞれ採用している.
[1] B. West Geoffery, A General Model for the Origin of Allometric Scaling Laws in Biology, SCIENCE VOL.276 (1997)
[2] A.Takamatsu, A.Takaba, E.Takizawa, G.Environment-dependent morphology in plassmodium of true slime mold Physarum polycephalum and a network growth model.J.Theor. Biol., 256, 29-44 (2008)
[3] 五味拓真, 真正粘菌変形体における酸素消費量の形状依存性, 早稲田大学大学院 先進理工学研究科 電気・情報生命専攻 修士論文 (2008)
[4] 平井友, 真正粘菌変形体におけるアロメトリックスケール則, 早稲田大学大学院 先進理工学研究科 電気・情報生命専攻 修士論文 (2009)
[5] K. Schmidt-Nielesen, スケーリング:動物設計論 動物の大きさは何で決まるのか 64-84, コロナ社 (1998)
2012年度
真正粘菌変形体における厚み振動のシミュレーション 阪田 佳亮
私は真正粘菌変形体における厚み振動現象を数理的に検証を行っている。真正粘菌変形体は単細胞生物であり、細胞内には複数の核が存在している。細胞内では原形質が往復流動している。また、培地の条件に依って異なる振舞いが報告されている。例えば、形状に着目すると、餌となるオートミールを含む培地ではシート状に成長し、忌避物質であるKClを含む培地では樹状に成長する。
また、細胞の厚みは振動している。この厚み振動は粘菌の形状によって異なり、シート状に成長した粘菌では螺旋状に伝播し、相対的に高い振動数で振動する。樹状に成長した粘菌では同心円状に伝播し、相対的に低い振動数で振動する。神経系を持たない単細胞生物が、このように異なる振る舞いを見せることは非常に興味深い事である。[1]
この厚み振動は、アクチン繊維の構造が関わっていると考えられている。アクチン繊維が網目状に結合すると、細胞内の原形質がその部位から押し出される。その結果、細胞の厚みは減少する。また、アクチン繊維の網目状の結合が分離すると、周囲から原形質が流入し、細胞の厚みは増加する。アクチン繊維同士の結合・分離には、Ca2+、ATP、cAMP、NADHなどの化学物質が関わっており、これらの化学物質は細胞の厚み振動を同じく振動していることが報告されている。[2]
しかし、多くの化学物質が関わっている点などを考慮すると、その振る舞いを「正確に」数理的に記述することは困難である。そこで、厚みという指標を局所的な原形質量として捉え、その振動を局所的に非線形振動子として抽象化する。そして、非線形振動子間では相互作用を及ぼしあう結合振動子系を考えている。また、このままでは、単に現象を抽象化しただけの数理モデルに過ぎないため、そこに厚み振動の特徴的な点を加えている。具体的には、系全体で原形質量が保存されているということである。
これらのことを考慮して実験を行い、現実系に近い結果を得られる数理モデルを得ることを目指している。更に、得られた結果を現実系にフィードバックし、厚み振動現象の本質について考察を行う。
[1] 高松敦子, “生きた細胞による結合振動子系の構築-複雑な振舞いをする生物観察の構成的アプローチ-“, 複雑系叢書6. コンプレックス・ダイナミックスの挑戦, 早稲田大学複雑系高等学術研究所, 共立出版, 2006
[2] 上田哲男, 粘菌細胞の知覚行動:非平衡化学ダイナミクスによるインテリジェンスの自己組織化, 日本機械学會誌97(902), 34-37, 1994
シアノバクテリアの運動解析 田中 慶太
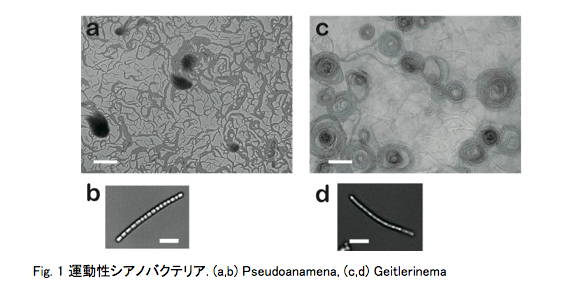
生物はしばしば“群れ”をなす。人間や哺乳類では複数の個体が集まってコミュニティーをなすことがあるが、魚や鳥などの動物も複数の個体が集まって群れをなして移動を行うことが知られている。そして、その移動の様子は単体でいる時と群れをなしている時では明らかに挙動が異なることがある。
興味深いことに、この“群れ”の習性はシアノバクテリア(Fig.1)でも確認することができる。バクテリアは一個体では単純で無秩序な動きをするが、複数存在するとコロニーを形成し複雑な動的パターンを形成する。つまり“群れ”は生物の本能からの行動であるといえるだろう。また、自動車はある程度密度が大きくなるとすぐ渋滞を起こし、全体の移動速度が低下するのに対し、生物は一般的に群れを形成し、密度が大きくなったとしても全体の速度が大きく低下することはない。ここにも生物特有の“群れ”の傾向がみられるのではないだろうか。そして、その傾向はシアノバクテリアにも適応できるのだろうか。本研究ではこの交通流の問題も交えて研究を行っている。
具体的に、本研究ではこのシアノバクテリアの“群れ”の運動を解析し、モデル化することまでを見据えて研究を進めている。まずシアノバクテリアの動画に対してPIV解析を行い、次にシアノバクテリアのモデル化の前段階としてアリの行進のモデル化を行った。また、それぞれの指標として交通流と比較するために速度と流量の二つを用いている。
(※PIV:Particle Image Velocimetory 流体速度解析のこと)
(※流量とは、一定時間の内にどれほどの個体が通過したかを表す指標である)
マイクロ流体デバイスを用いた海洋微生物の現場生菌数測定 および水産物に含まれるウイルスの検出・定量 鳴澤 良友

海洋環境における微生物相の構成に関する研究は現場遺伝子解析装置の開発などにより急速に進展している. その一方で,現場における微生物活性の指標としての生菌数計測については従来手法が用いられている. 現在のサンプル採取に依存した分析手法では外来微生物が混入する可能性や,水圧や温度等の変化に伴い活性が変化する可能性がある. 本研究ではこのような現行手法の課題を解決するため, 深海微生物の生菌数を現場で計測できる装置の開発を目指している. 海中探査機に搭載する測定機器は小型であることが求められるため, 小型で高度な流体操作が可能なマイクロ流体デバイスを応用する. マイクロチャンバを用いた生菌数計測法は,様々な体積を有する複数の円柱状チャンバに微生物を含む試液を封入・培養するものである。ある細胞密度のサンプルを封入した時にチャンバに微生物が一細胞も入らない確率は理論的に求めることができるため, 逆に増殖が生じなかったチャンバ割合からMPN(Most Probable Number)法と同様の確率的手法で試液中の生菌数密度を推定することができる[1]. ここでは直径がそれぞれ640, 320及び160µmで深さが40µmの大中小3種の円柱状チャンバを持つマイクロデバイスに7.13×104細胞/mlの酵母サンプルを封入した(Fig. A). これを培養後、無菌チャンバ数から算出した密度と元のサンプル密度との比較を行った. 結果として7.13×104細胞/mlの酵母サンプルの場合、誤差は最大で6.17%であった (Fig. B)。今後、微生物の封入方法や各チャンバにおける増殖の有無の検出法を検討・評価することで、マイクロチャンバを用いたMPN法による現場生菌数測定に展開することができる.
更に現在はマイクロチャンバデバイスを用いたMPN法を応用し, カキなどの水産物に含まれるカキヘルペスウイルスやノロウイルスなどのDNA・RNAの実験室環境での検出・定量を行おうとしている. 微生物の生菌数測定法における微生物は水産物サンプルから抽出したDNAやRNAに相当し, 微生物の培養によるチャンバに封入されたかどうかの検出はDNAやRNAの増幅に相当する. チャンバ内におけるDNA増幅をどう実現するかが課題となる.
外部研究先 東京大学生産技術研究所 藤井輝夫研究室 福場辰洋
[1] William G. Cochran, Biometrics, 6(1950) pp.105-116
[2] V. Flemming and I. Kjeld, Appl. Environ. Microbiol., 63 (1998) pp. 1700-1707
2013年度
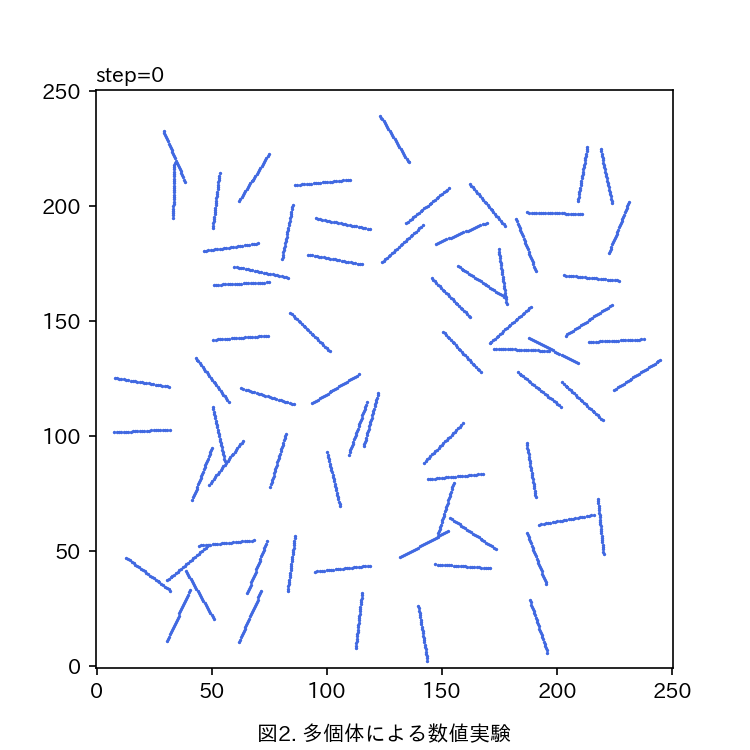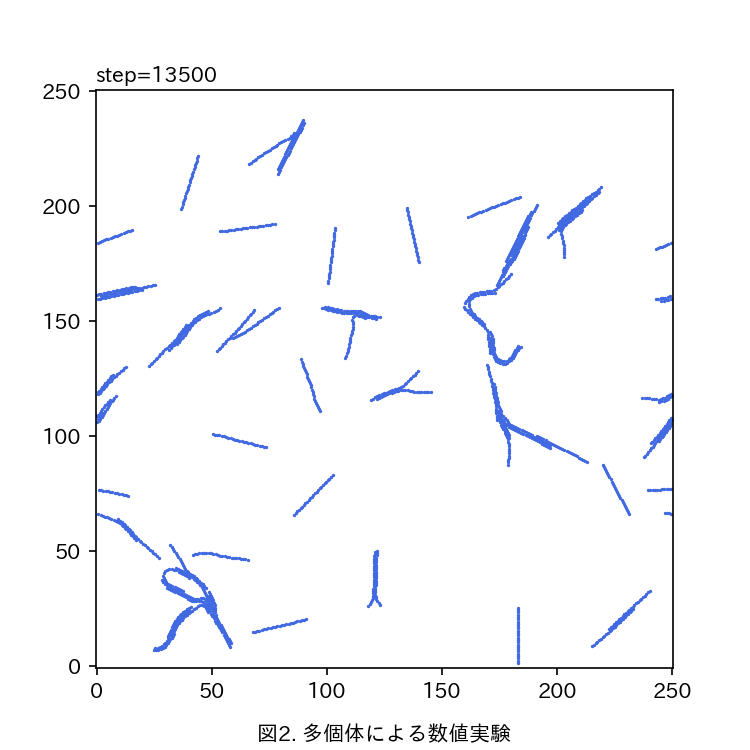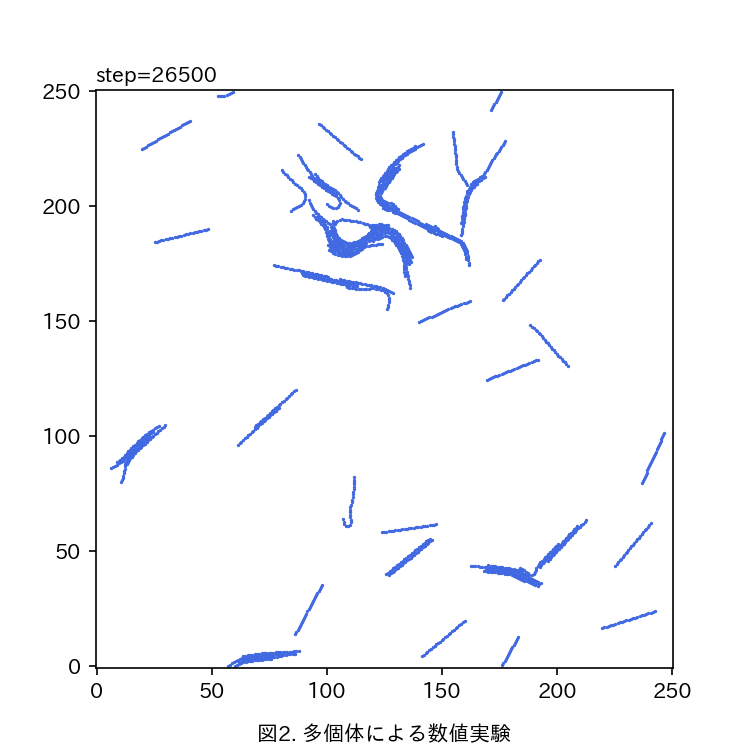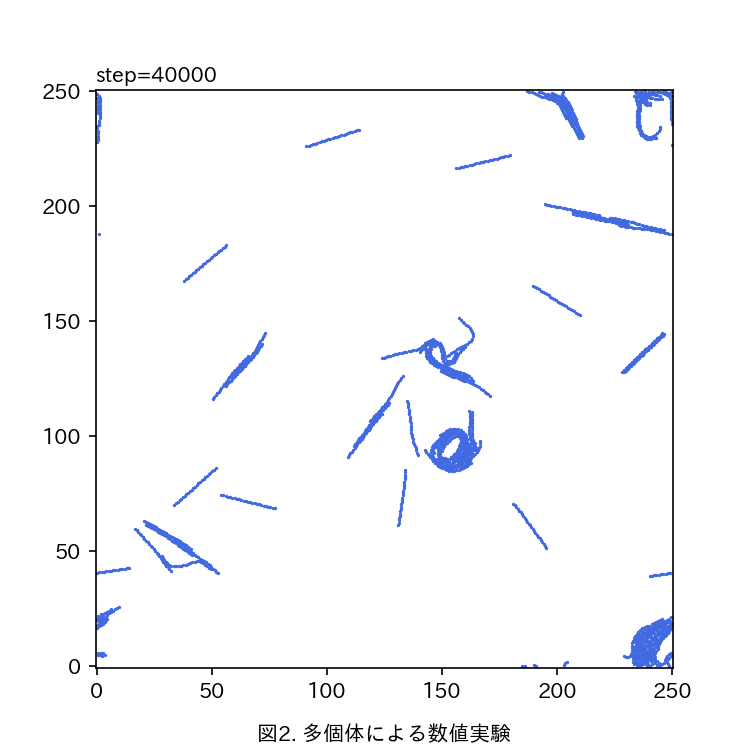
真正粘菌変形体の作る輸送管ネットワークの成長 の定量とモデル化 青山 隼也
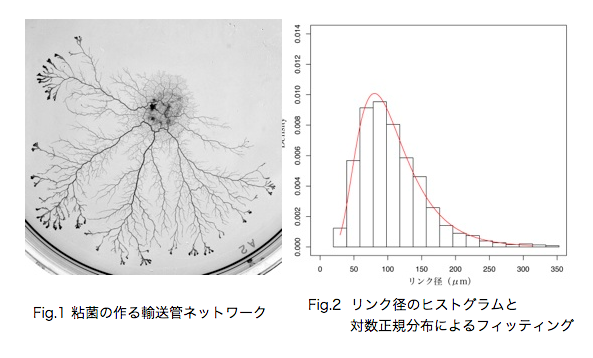
真正粘菌変形体をモデル生物として、生物の形成する輸送管ネットワークの定量的な解析を行う。真正粘菌変形体は多核の単細胞生物で、その構造は比較的単純な生き物である。数十μm〜数mまで多様な大きさで存在することが可能で、輸送管ネットワークを形成し細胞中の栄養分や酸素の運搬を行っている。また、真正粘菌変形体は単純な構造であるにも関わらず環境条件によって形態を大きく変化させたりといった複雑な情報処理が必要と思われるような生物らしい振る舞いを示す。情報の伝達も輸送管を通して行われると考えると、輸送管ネットワークが情報処理などの生物らしさを粘菌に与えていると言える。私はこの粘菌の輸送管ネットワークに着目し、様々な環境条件で成長過程を観察している。輸送管ネットワークの分岐と分岐点をつなぐリンク部分をひとつの単位として、この部分の太さ(リンク径)が成長とともにどのように変化していくのかを調べたところ、リンク径は対数正規分布をなしていると言えそうである。対数正規分布は様々な要素が複雑につながり合った系(複雑系)においてよく見られる確率分布であり、過去からの様々な確率的要因の積み重ねとして生じるような乗算過程において現れる。このことから、粘菌は原形質流動を繰り返すことによって輸送管径を変化させ、そこに確率的なゆらぎが加わることで、対数正規分布として観測されるということができる。 また、観察結果から対数正規分布のパラメータを推定することで成長過程による時間発展を定量的に評価できる。 いずれのサンプルにおいても分布の形状がほぼ同じであることから輸送管の成長の背後には一貫した法則性があることが分かる。 いずれの環境においても対数正規分布をなすという点では共通であるが、パラメータの変化の仕方には違いが見られる。現在どのようなメカニズムで対数正規分布が現れるのかを考え、輸送管成長過程の数理モデルとしての表現を模索中である。
2014年度
シアノバクテリアにおける個体間の相互作用解析 工藤 亮太
多くの生物は群れを形成して生活している。哺乳類や鳥類、魚類などが群れをなしているのは想像に難くないであろう。群れをなす性質は、その群れが有利になるために形成されると考えられる。つまり、群れの形成はその種の保存を助けているのである。
群れを形成する性質はシアノバクテリアにおいても同様である。シアノバクテリアは複数の個体が寄り集まって種々の形状の集団(以下コロニー)を形成して運動を行っており、その形状は一定ではなくコロニーのパターン間で動的に変化する。コロニーが形成され、形状が変化する要因として個体間での相互作用があげられる。個体間での相互作用によって個体同士が意図的に寄り集まり、その積み重ねによってコロニーが形成されるのである。通常、群れ形成のモデルで用いられる相互作用は、個体同士の距離のみに依存したポテンシャル関数を採用している。しかしシアノバクテリアは、離れて運動していた個体同士が急激に引き寄せ合う、個体同士が近づいた際に進行方向を変えて接触を避ける、といった挙動を示すため、距離のみに依存したポテンシャル関数ではうまく表すことが出来ない。そこで、個体同士が引き寄せ合う、反発するといった挙動を観察するためにシアノバクテリアを連続で画像撮影、解析し、個体間距離に加えて、それぞれの個体の運動速度や二個体の進行方向が成す角度などを計測する。これらのデータを解析することで、個体間に働く相互作用を解析する。
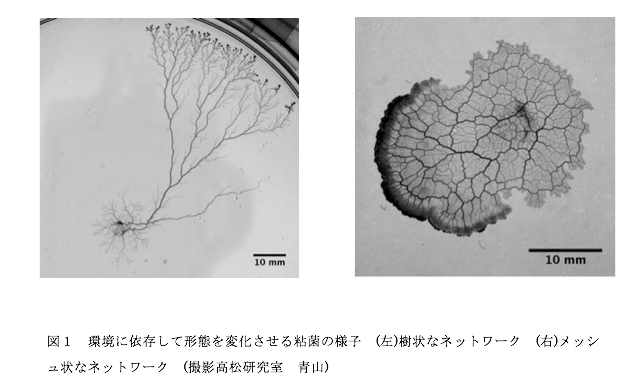
真正粘菌変形体における管径成長の環境依存性 山口 高輝
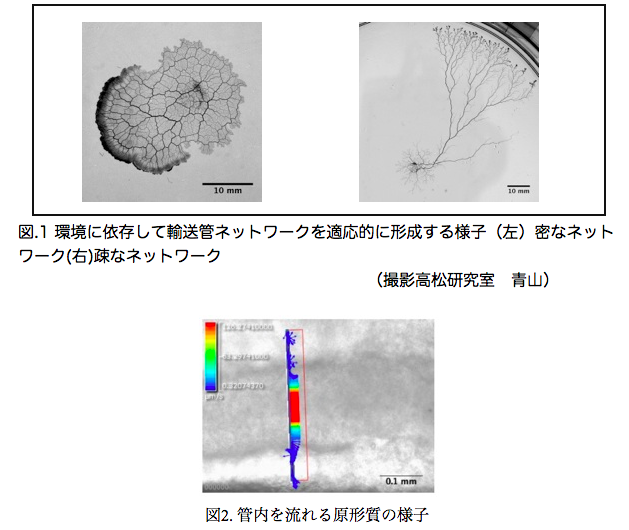
真正粘菌変形体(以下、粘菌と呼ぶ)は管状の輸送管ネットワークを形成する。管内では原形質往復流動を通して栄養分や酸素を粘菌全体に運搬する。この輸送管ネットワークの形態は培地の寒天濃度、化学物質の濃度に依存して変化することが知られている。固い培地や栄養分が豊富な環境では細い輸送管で構成される密なネットワークを形成する。柔らかい培地や忌避物質を含む環境では太い輸送管で構成される疎なネットワークを形成する [1]。このように単細胞生物でありながら環境に依存して形態変化させ、最適なネットワークを形成する適応性を備えている[2](図.1)。輸送管の管径は管中を流れる原形質の流量に応じて変化する性質をもつ。すなわち、流量が多いほど管径が大きく、少ないほど小さくなる(図.2)。この管径と原形質の流量の関係はヒルの式を用いた管径成長方程式で表せる[3]。
本研究では環境依存性を調べるため、誘引環境、中間環境、忌避環境の3つの異なる培地環境を用意した。培地上に設置した鋳型に粘菌を配置し、輸送管の成長過程を撮影した。撮影した動画から原形質の流速、管径、流量を算出した。得られたデータから管径成長方程式におけるパラメータを推定することで環境依存性について評価した
実験の結果、管径成長過程において誘引環境と忌避環境では原形質の流速、管径、流量の時間変化に差異が観察された。誘引環境では管に流れる流量に応じて緩やかに管径が成長する。忌避環境では流速がある程度大きく流れ始めてから勢いよく管径が成長する。以上の流量と管径成長の関係から、成長方程式のパラメータを推定した。そのうち減衰係数には環境毎に有意差が見出せたが、成長項の収束値、ヒル係数、規格化に用いたパラメータについては見出せなかった。このことから、管形成長のダイナミクスを明らかにするためにさらに分析を進める予定である。
[1] M. Ito, R. Okamoto, and A. Takamatsu (2011) J. Phys. Soc. Jpn., 80, 074801
[2] A. Takamatsu et al. (2009) J. Theor. Biol., 256, 29-44
[3] A. Tero et al. (2007) J. Theor. Biol., 244, 553-567
真正粘菌変形体における培地環境と振動数の関係 西川 哲平
真正粘菌変形体は多核の単細胞生物であり、数十μmから1m程度まで成長する。厚み振動における原形質流動によって身長成長を行う。 さらに培養する培地に適応し、形状を変化させる。粘菌の餌であるOatmealを含む培地や硬い培地などの誘引環境だとメッシュ状、KClを含む忌避環境や柔らかい培地だと樹状に変化する。脳や神経系を持たない粘菌はどのようにして培地環境を認識しているのだろうか。本研究では粘菌が環境を認識する際に、何を基準にして培地を判断しているかを解明することを目的とし、培地に占める面積比率を角振動数に着目して解析を行った。シャーレを真ん中で粘菌を培養した時の成長過程を画像解析した。結果として寒天濃度、Oatmeal、KClの条件の中でOatmealが粘菌の培地認識に大きく影響していることがわかった。今後の課題として面積比率に差が生じたところに着目してさらに詳しく解析を行う必要があることが挙げられる。また、面積比率や角振動数という指標とは別に位相についての解析を行うと新たな結果がでると思われる。
2015年度
粘菌アルゴリズムにおける 環境の時間変化を考慮したネットワーク設計 小野 恭吾
本研究では、真正粘菌変形体(以下、粘菌と呼称する)という生物から着想を得たアルゴリズム[1]を用いて、環境の時間変化を考慮したネットワークにおける最適化を考える.すなわち、環境の時間変化にさらされて段階的に建設されるネットワークと、初めから計画されており同時に建設された際のネットワークを区別して考え、シミュレーションにより比較をする.
私たちが普段利用する交通網、電力網などのネットワークについて考えてみてほしい.いかなるネットワークも、取り巻く環境は時々刻々と変化している.電力網を例にすれば、既に電線の通っている街に新しい発電所や住宅地が出来ることがある.この際、段階的に最適化されたネットワークは、初めからその環境になることが計画されていたときに出来上がるネットワークとは異なると考えられる.これは段階的に最適化されたネットワークは、既にあるネットワークを考慮して新たにネットワークを形成するが、初めから計画されていた環境であればネットワークも計画の通りに形成するからであると考える.
粘菌は多核の単細胞生物であり、数十mから1m程度にまで成長し、平面上に輸送管を張り巡らせるようにネットワークを形成して進む.輸送管内では核、ミトコンドリア、細胞質などを含む原形質の往復流動を行い、栄養分や酸素の運搬をする.輸送管内の管径は、管内を流れる原形質の流量に応じて変化する性質を持つ.すなわち、流量が大きい管径は太く成長し、小さい管径は細く減衰する. 粘菌は、情報を処理する中枢を持たないのにも関わらず、このような適応過程を経て効率の良いネットワークを形成する.先行研究では、迷路に粘菌を敷き詰めスタートとゴールの位置に餌を与えると、最短経路を繋ぐネットワークを形成することも示された[2].
現在は、環境の時間変化を考慮して段階的に建設されたネットワークと、同時に建設されたネットワークを脆弱性やエネルギー損失などの観点から評価を行っている.今後は、実際の道路網情報に当てはめて評価していきたいと考えている.
[1] Tero, A., Kobayashi, R., Nakagaki, T., 2007. A mathematical model for adaptive transport network in path finding by true slime mold. J. Theor. Biol. 244, 553-564.
[2] Nakagaki, T., Yamada, H., Toth, A., 2000. Maze solving by an amoeboid organism.Nature 407, 470.
真正粘菌変形体の培地環境に依存するネットワーク中心性について 辻 喬

真正粘菌変形体は多核の単細胞生物であり成長する際に管状の輸送管ネットワークを形成する。粘菌は脳などの中枢神経系を持たないにもかかわらず、ネットワークの形態を環境に合わせて変化させることが知られている。
本研究では環境に依存するネットワーク形態の生物学的機能を調べるために、媒介中心性というネットワーク内の重要な部分を評価するための指標に着目した。粘菌ネットワークの管を枝、管の交差点を分岐点として、実際に異なる環境における粘菌ネットワーク媒介中心性を算出した。
解析結果から忌避的環境と誘引的環境では重要な管が異なることが示唆された。粘菌にとって忌避的環境では放射方向に伸びた管が重要であると考えられるのに対し、誘引的環境では加えてそれらをつなぐような円周方向の管も重要になってくると考えられる。
光刺激状況下における真正粘菌変形体のネットワーク形成 豊田 悟史
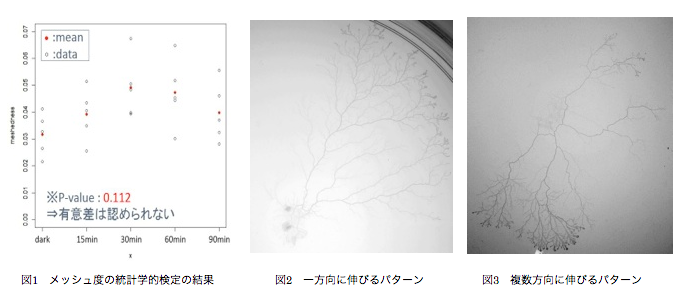
本研究の目的は、時間変化する光刺激に対する粘菌の適応の仕組みを、ネットワーク構造という側面から解明することである。粘菌には光を避ける性質があることが知られている。そこでネットワークの成長過程で周期の異なる光刺激を加えた場合にネットワーク構造が変化すると仮定し、実験および解析を行う。具体的な手法としては、まず周期の異なる光刺激を与える実験を行い、粘菌ネットワークの時間変化を撮影する。次に撮影した写真に画像処理を施し、管の長さや太さなどのネットワークの特徴量を抽出する。抽出されたデータを元に、そのネットワーク構造の分析を行う。分析には「つながり方」に注目するネットワーク理論による分析や、形に着目する幾何学的な分析を利用する。
実験結果に基づき、ネットワークの粗密性を評価する指標である「メッシュ度」を用いて、ネットワークの枝数と頂点数の比率に差が見られるかどうかを解析する。統計学的検定により確認すると、有意差は認められないという結果となった(図1)。これは、条件間で枝数と頂点数の比率に違いはほぼ見られないということを示唆している。
一方で、刺激の無い条件ではおおよそ一方向にネットワークが成長していく様子が見られるのに対し(図2)、ある周期の条件では複数の方向にネットワークを伸ばしていく様子が観察された(図3)。粘菌がこのようなネットワーク形成を行う理由として、次のことが仮説として考えられる。たとえば一方向にのみ成長している場合だと、その部分が破壊されることで生存の危機にさらされてしまう。対して、複数方向にネットワークを伸ばしておけば、一部が破壊されても他の部分は生存できる。このように、粘菌は不安定な環境におかれると、生存確率を上げるために「保険」としてのネットワークを形成する可能性がある。さらに先ほどのメッシュ度についての結果を併せて考えると、枝数と頂点数の比率を大きく変えずに、つまり「枝を増やすためのコスト」を抑えつつネットワークの形態を変化させることで、生存確率を上げているとも考えられる。
今後は以上の結果と仮説について定量的な分析を進めるとともに、どのような機構が存在すればこのような応答が実現できるのかを、モデルを利用した理論的なアプローチも試みる予定である。
生物輸送管ネットワークの適応過程に着想した ネットワーク最適化 渡辺 晋
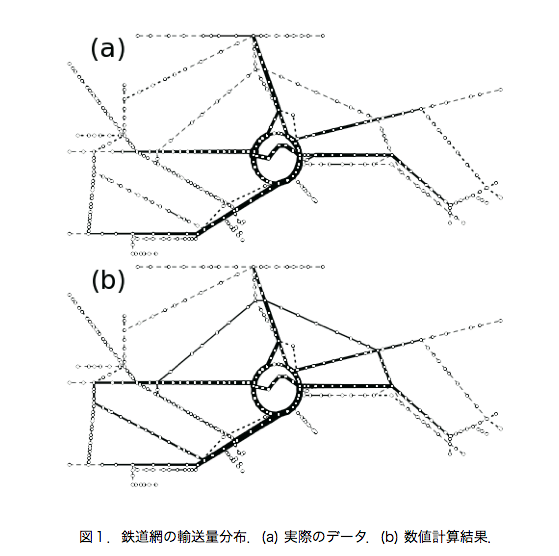
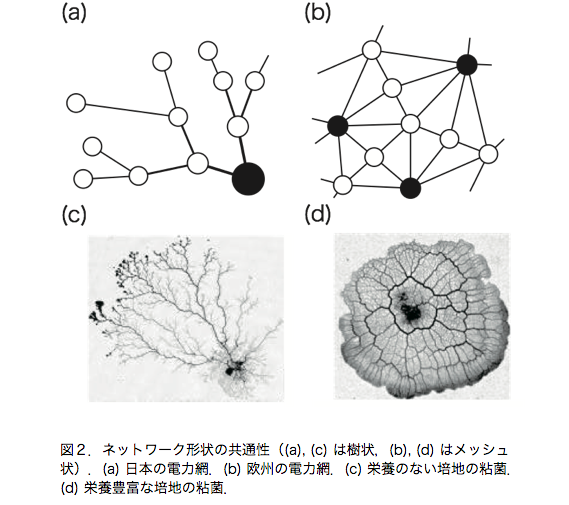
生物の適応的な振舞いに着目した手法を用いたネットワーク設計について研究を行っている.具体的には真正粘菌変形体が形成する輸送管ネットワークをモデルにしたアルゴリズム[1]を応用して鉄道網や電力網のネットワーク設計に取り組んでいる.
真正粘菌変形体(以下,粘菌と呼ぶ)は多核単細胞生物であり,その大きさが10µm-1mほどにも成長する.このように大きな細胞体を維持するために体内に形成した輸送管を通して原形質流動を行うことで酸素や栄養素などを全身に行き渡らせている.そして,原形質流動が盛んな管ほど太く成長しやすく,乏しい管は減衰・消滅するという適応的な特徴が見られる.このような適応過程を経ることで粘菌は効率の良いネットワークを形成している.例えば迷路に粘菌を敷き詰め入口と出口に餌場を設けると粘菌は適応過程を経て入口と出口を結ぶ最短経路に管を残す[2].また広がった粘菌に複数の餌場を与えると輸送効率や断線保障性(任意の管が切れてもネットワーク全体が繋がっている性質)の優れた輸送管ネットワークを形成する[3].
このような粘菌の適応過程に着想を得たアルゴリズムを応用して鉄道網の輸送量設計に取り組んできた[4].ネットワークを路線図の形状に固定して,餌の量を駅の利用者数に比例させた条件で輸送量分布を計算した.その結果,図1のように実際の鉄道路線の利用状況によく似た輸送量分布を得ることができた.また得られた分布にネットワークの維持コストや利便性,耐障害性に相当する評価関数を用いて計算したところ,評価関数値のバランスが取れた分布になっていることが分かった.人間が意図的に作ってきた鉄道網における輸送量分布が生物的な過程を経ている可能性があり,結果としてバランスの良いものになっているという興味深い結果が得られた.
現在,この手法を応用して電力網設計に取り組んでいる.図2のように粘菌の輸送管ネットワークは栄養のない培地では樹状に,栄養豊富な培地ではメッシュ状に広がることが知られている[5].また電力網においても日本では原子力発電所や火力発電所を中心に樹状にネットワークを形成しており,欧州では豊富な自然エネルギー発電所が点在しておりメッシュ状に発達している[6].このような共通点を持つ粘菌をモデルにしたアルゴリズムを用いることで電力系の問題を考慮した電力網ネットワーク設計を目指す.既存のネットワーク設計では需要量を一定としていたが,電力網では需要量の時間変動を想定して送配電線ネットワーク設計を考える必要がある.例えば,夏期や冬期に需要量が増加し,一日の内でも需要家の活動時間帯に需要量が増加する.さらに活動時間帯は各需要家によって異なる.本研究では,発電所1基と需要家2軒から構成される単純なネットワークについて解析を行う.そして,需要家では需要量が変動し,その変動の位相差をパラメータとして与えネットワーク形態を計算する.
[1] Tero, A., Kobayashi, R., Nakagaki, T., 2007. A mathematical model for adaptive transport network in path finding by true slime mold. J. Theor. Biol. 244, 553-564.
[2] Nakagaki, T., Yamada, H., Toth, A., 2000. Maze solving by an amoeboid organism. Nature 407, 470.
[3] Tero, A., et al., 2010. Rules for biologically inspired adaptive network design. Science 327, 439-442.
[4] Watanabe S., Tero A., Takamatsu A., and Nakagaki T., 2011. Traffic optimization in railroad networks using an algorithm mimicking an amoeba-like organism, Physarum plasmodium. Biosystems 105, 225-232.
[5] Takamatsu, A., Takaba, E., Takizawa, G., 2009. Environment-dependent morphology in plasmodium of true slime mold Physarum polycephalum and a network growth model. J. Theor. Biol. 256, 29-44.
[6] 経済産業省資源エネルギー庁電力基盤整備課, 「電力系統の構成及び運用に関する研究会」報告書, 平成19年4月17日公表
シアノバクテリアの運動機構の解明 齋藤 拓
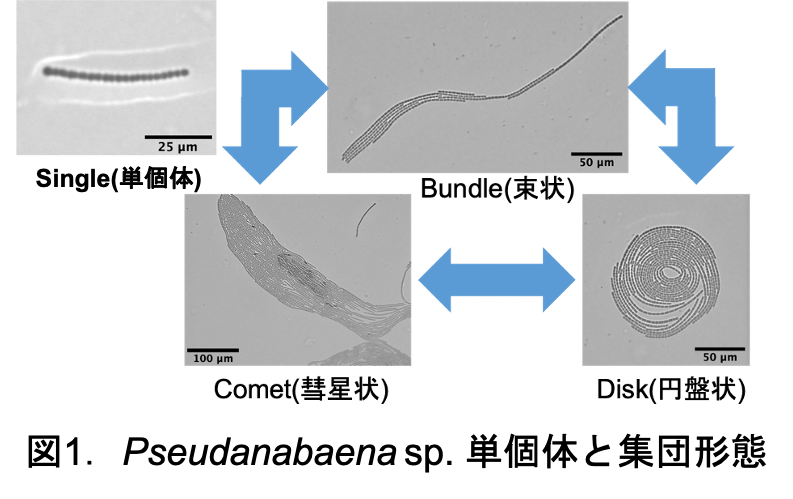
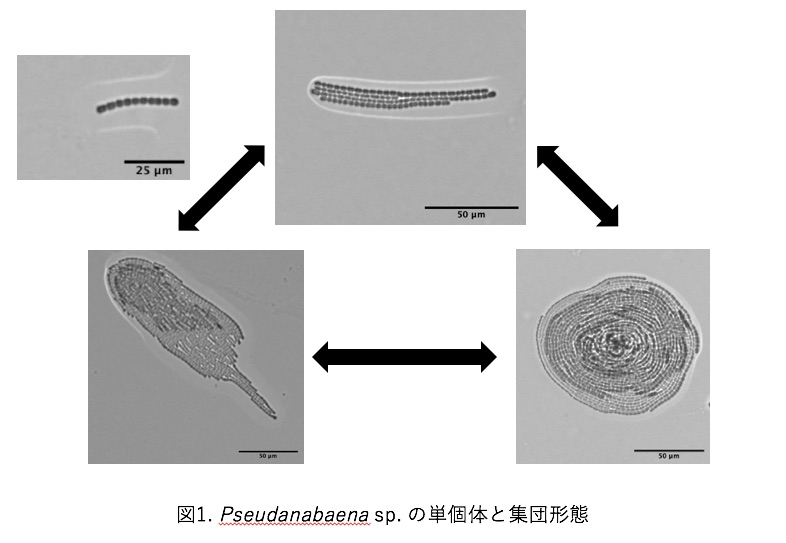
シアノバクテリアの一種であるPseudanabaena sp. ILC545は、ユレモ目に属する糸状性のシアノバクテリアである。4μm程度の長さの栄養細胞が連なって1個体を形成する。1個体が単体で運動を行う際は主に長軸方向に運動し、個体同士が集合することで群れを形成する。群れを形成することによってシアノバクテリアの運動は劇的に変化する。代表的な群れとして個体が積層して速度も大きい彗星状の群れであるcomet、同じ場所に留まり、回転運動をする円盤状のdisk、この2種類が存在する。このようにPseudanabaenaの特徴の一つとして、群れを形成するとこによって1個体では成し得ない運動をすることが挙げられる。しかし群れ形成の要因は未だ解明されていない。仮説として、群れ形成の要因として考えられるものの一つにシアノバクテリアの運動機構が挙げられる。しかし、Pseudanabaenaの運動機構についても未だ解明されていない。私は群れ形成の要因解明のボトムアップ研究として、シアノバクテリアの運動機構の解明を研究目的に実験を行っている。
研究方法として現在2つのアプローチを行っている。1つ目は観察である。高倍率かつ短間隔の動画を撮影し、シアノバクテリアの細かい運動を観察し、解析することで研究を行っている。解析方法としては、輝度値測定などを用いて運動機構の解明を目指している。2つ目は蛍光ビーズを用いた実験である。シアノバクテリアが運動を行う際に、周囲にいる小さいシアノバクテリアが影響を受けている様子が確認された[図1]。ここからシアノバクテリアは周囲の場に対して作用していると考えられるため、運動しているシアノバクテリアの周囲に蛍光ビーズを散布し、ビーズの運動を解析することにより場の相互作用の解明を行い、運動機構に対するヒントが得られないかと考えている。
血管形成におけるネットワーク解析 関根 渉

血管は体内で非常に大切な役割を担っている。組織細部まで血管が張り巡らされることで体の隅々まで酸素と栄養分が供給され、生命活動を保つことができている。図1を見ると、血管ネットワークは健康状態では秩序立っており、規則正しく形成されているように見える。しかし例えば腫瘍血管など異常状態になると、秩序は見られず場所によって密度が異なるように血管形成が行われている。[1]
そこで本研究ではネットワークの観点から健康な血管と異常状態の血管の違いを定量化することを目的とする。正常状態と異常状態を数理的に明確にすることで、血管治療における改善度指標として提案したいと考えている。また、指標としてはネットワーク全体のつながりを示す「平均頂点間距離」や網目状の指標である「メッシュ度」といった指標を用いる。例えば血管形成が促進され、より密なネットワークになれば平均頂点間距離は短く、メッシュ度は高くなるという関係である。 現在マウス網膜血管の画像を用いている.[2]正常血管の数理的定義を明らかにした上で、今後虚血性網膜症モデルマウスの網膜血管を用いて、改善度指標の提案に向けて解析を行っていく予定である。
[1]Yuval Gazit et al, Scale-Invariant Behavior and Vascular Network Formation in Normal and Tumor Tissue, Phys. Rev. Lett. 75, 2428,1995
[2] Fukushima Y et al, Sema3E-PlexinD1 signaling selectively suppresses disoriented angiogenesis in ischemic retinopathy in mice, J Clin Invest.121:1974-1985, 2011
2016年度
真正粘菌変形体の摂餌選択における集団意思決定 奥野 純平
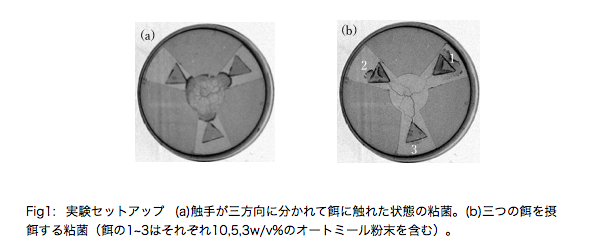
摂餌における意思決定は、すべての生物にとって重要な生存戦略であり、これは脳を持たない単細胞生物においても例外ではない。本研究の対象生物である真正粘菌変形体(以下、粘菌と呼ぶ。)は、多核の単細胞生物であり、その巨大な細胞を触手のように伸ばすことで餌の探索、摂餌を行う。先行研究において、粘菌は摂餌の際に、有害な光刺激と餌の利得とのトレードオフや、意思決定の速度と正確性とのトレードオフを行うことが知られている[1,2]。これらの先行研究では、粘菌の意思決定における最終段階に着目しているが、実際には粘菌は多方向に触手を伸ばし、複数の餌を同時に得ることができる。そこで本研究では、多方向に触手を伸ばした粘菌の意思決定のプロセスに着目した。
実験では、三つのブロック状の餌を、三方向に伸びた粘菌の触手に触れるように設置した(Fig.1a)。培養し始めてから3〜12時間後には、触手がそれぞれの餌を選択的に摂餌していく様子が観察された(Fig.1b)。
餌の濃度と触手の成長速度との関係を、餌を覆い尽くした時間や餌の滞在時間から解析を行った。その結果、三つの餌の質が同じ場合は、餌の選択順が触手の進展速度に依存することが示唆された。対して、三つの餌の質が異なる場合は、餌の質と進展速度の複合的な要因に依存することが示唆された。
[1] Latty, Tanya, and Madeleine Beekman., 2010. “Food quality and the risk of light exposure affect patch‐choice decisions in the slime mold Physarum polycephalum.” Ecology, 91.1: 22-27.
[2] Latty, Tanya, and Madeleine Beekman., 2011. “Speed–accuracy trade-offs during foraging decisions in the acellular slime mould Physarum polycephalum.” Proceedings of the Royal Society of London B: Biological Sciences, 278.1705: 539-545.
ゼブラフィッシュの分子時計の光応答機構 吉次 なぎ
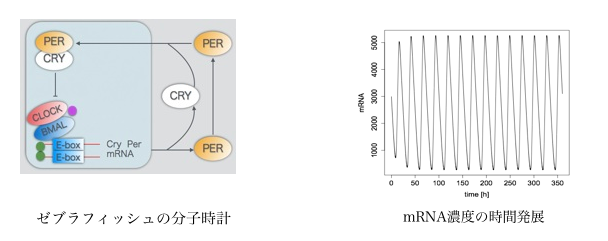
生物は様々なリズム現象を示す. 心臓の鼓動, 種子形成のリズム, 蛍の集団発光などがその代表的な例である. リズム現象の機序を解明するために, 分子生物学や生理学的アプローチが取られる一方で, 数学的手法を用いることも有効である. 現象を記述する数理モデルを構築し解析することで, 現象が起こる条件を正確に特定することや未知の現象を予言することができる[1].
多くの生物が概日リズムを持っている. 概日リズムとは生体内のおおよそ24時間のリズムのことで, 生物種や個体によって概日周期は異なり, 人間の概日周期は平均24.11[h]と言われている. 多くの生物で24時間から多少のズレがあるが, 光入力などによって調整されることが知られている[2].
ゼブラフィッシュ(Danio rerio) は飼育, 観察の面で優れ, 光を受容する網膜の構造がヒトとを近いため, 概日リズム研究のためのモデル生物として注目されている[3]. 多くの脊椎動物では光入力が網膜を通して体全体の概日リズムを調節するが, ゼブラフィッシュはこれに加えて細胞が直接光入力を受け取ることができ, 光入力に対して鋭敏に反応することが知られている[4]. 本研究では, ゼブラフィッシュの1細胞に着目し, 細胞内の分子時計が光によって調節される仕組み, 条件を数理モデルを用いて探ることを目的としている. まず光入力がない状態(フリーラーニング)での1細胞の分子時計を数理モデルで再現し, その後光入力を考慮したモデルを構築する方法を考察する.
[1] 三村 昌泰, 竹内 康博, 森田 善久 (2011), 生物リズムと力学系, 共立出版.
[2] 海老原 史樹文, 吉村 隆 (2012), 時間生物学, 化学同人.
[3] Emran Farida, John E. Dowling(2014), “Circadian Rhythms and Vision in Zebrafish.” The Retina and Circadian Rhythms, Springer New York.
[4] Tamai T. K. Cara, A. J. Whitmore D(2005), “Zebrafish circadian clocks: cells that see light.” Biochemical Society Transactions.
シアノバクテリアが形成するコロニーパターンと環境の関係 佐藤 隆久

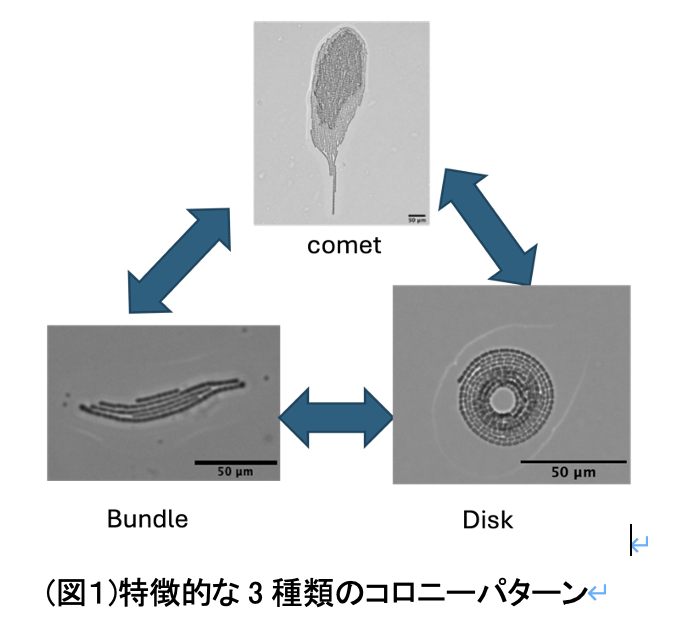
シアノバクテリアの一種であるPseudanabaena sp.ILC545は粘液を分泌しながら寒天培地上を滑走運動する.さらに複数の個体が集まると,3種類のコロニーを形成することが知られている.2個体以上が集まってできる束状のbundle,運動速度が速い彗星状のcomet,その場に留まり回転運動をする円盤状のdiskである.これらのコロニーは時間と共に動的に形状変化し,パターンを形成する.以上のようにPseudanabaena sp.ILC545が3つのコロニーパターンを形成することは知られているが,各コロニーの生物学的メリットは明らかにされていない.
そこで本研究の目的はPseudanabaena sp.ILC545の各コロニーの生物学的メリットを提言することとする.図1にシアノバクテリアの培養の様子を示す.図で示した4つの培地は寒天濃度を変え,右に行くほど固い培地になっている.するとどれも6日間経過しているにも関わらず,生息範囲の広がりに差があることが確認できる.また図2に標準の培地である寒天濃度1.5w/v%の拡大図を,図3に用意した中で最も固い培地である寒天濃度5w/v%の拡大図をそれぞれ示す.すると出現しているコロニー形態にも差があるように思われる.
そこで私は寒天濃度がコロニーの形成に影響を及ぼし,ひいては生息範囲拡大にも影響しているのではないかと仮説を立てた.具体的には寒天濃度の低い培地では運動速度の速いcometが多く出現するため生息範囲を速く広げることができるのではないかと考えている.現在は培地環境を変化させ,一日毎に各コロニーの出現割合を確認する実験を行い比較している.今後は寒天培地中の必須塩濃度も変化させることで,形成されるコロニーパターンの違いについて総合的に考察し,形成要因について提言することを目指す.
アリの協同運搬における行動の多様性の効果 宇都宮 祥平

アリが一個体では運搬できない大型のエサを発見した際には大まかに2種類の運搬方法を行う.それは仲間のアリと協同運搬をする方法(図1)と,大型のエサを一個体で運べるように解体して運搬する方法である[1].本研究では前者の協同運搬を扱う.協同運搬では全てのアリが巣の方向にエサを運搬するのではなく,巣とは反対方向にエサを運搬しようとするアリが確認されている.この巣とは別方向に運搬するアリが協同運搬においてどのような役割を持つかを解明することが本研究の目的である.
実験観察より,次の二つの協同運搬における特徴を定量化した.一つ目は,協同運搬の運搬速度が個体数の増加により変化しない場合でも,障害物を通過する際の通過時間が個体数の増加に伴って短くなることである.二つ目は,一個体による運搬でも巣とは別方向に運搬するアリが存在し,個体ごとの運搬方向にばらつきがあることである.これらの実験結果より「アリの協同運搬において,個体ごとの行動のばらつきが障害物通過を効率化する」という仮説が立てられる.この仮説を検証するために,本研究ではアリがエサを牽引する力学モデルを構築した.
このモデルではエサを運搬するアリが認識している巣の方向にゆらぎを与えることで,個体ごとの行動のばらつきを表した.このゆらぎがある場合とない場合でシミュレーションを比較することで仮説を検証する.シミュレーション結果より,個体ごとの行動にばらつきがある場合に障害物の通過時間が個体数の増加に伴って短くなることが確認できた.したがって,巣とは別方向に運搬するアリが障害物通過を効率化していることが示唆される.
[1] 伊藤文紀 (2009)「アリ類における大型餌の運搬」『昆虫と自然』44, 15-19
2017年度
粘菌ネットワーク形成を利用した 道路網上の縄張り境界の解析 佐藤 滉大
東日本大震災をはじめ、過去の自然災害では、避難所に到達できず多くの死者が出た。このことから避難所の最適区分の問題は多くの人命を救う上で重要である。今日、避難所の区分の幾何的な一般解はボロノイ図という手法で求めることができる。これは、避難所までの直線距離が最短となる領域で分割した図であるため、簡単に算出できることが利点である。しかし、道路網において、人の移動はネットワーク上に限定されるため、領域を直線距離で分割して得られる区分けは不正確となる欠点を持つ。そこで粘菌から着想を得た数理モデルである粘菌アルゴリズム[1]を用い、避難区分を考える。
ここで粘菌が作る区分けを粘菌勢力図と定義する。粘菌勢力図がどの程度経路最短かを表す指標としてネットワークボロノイ[2]を用いる。ネットワークボロノイはネットワークのリンクやノードを経路長に応じて分割した図である。本研究では、青梅市の道路網に、それらを適用し、避難区分を求める。最後に、得られた二つの避難区分を三つの指標、道路敷設コスト、移動時間と混雑度を用いて評価を行った。その結果、移動時間はネットワークボロノイとあまり変わらなかった。このことから粘菌勢力図がある程度経路最短であることが示唆される。混雑度とコストから、ネットワークボロノイにおける避難区分では高コストにも関わらず混雑度が高い結果となった。一方で粘菌勢力図の場合は低コストで混雑度が低い結果となった。このことから、粘菌勢力図による道路幅を用いることで混雑を回避できると示唆された。
[1]Tero, Atsushi, Ryo Kobayashi, and Toshiyuki Nakagaki. “A mathematical model for adaptive transport network in path finding by true slime mold.” Journal of theoretical biology 244.4 (2007): 553-564.
[2]Okabe, Atsuyuki, et al. “Generalized network Voronoi diagrams: Concepts, computational methods, and applications.” International Journal of Geographical Information Science 22.9 (2008): 965-994.
[3]Watanabe, Shin, et al. “Traffic optimization in railroad networks using an algorithm mimicking an amoeba-like organism, Physarum plasmodium.”Biosystems 105.3 (2011): 225-232.
ストレス応答を考慮した概日時計の数理モデル設計 染谷 洋輔
地球上の生物は、約24時間のリズムを刻む概日時計という機構を持つ。概日時計は身体的、精神的ストレスにより大きく乱れることが知られている[1]。本研究ではマウスの概日時計に与えるストレスの影響を再現する数理モデルを構築し、様々なストレス条件化における概日時計の挙動をシミュレーション可能にすることを目的とする。
本研究においてストレスを受けて概日リズムが乱れるのは、末梢時計間の調整因子として働くストレスホルモンの分泌量が増えることが原因であると仮定した。数理モデルとして光入力を受ける脳と、ストレスホルモンを分泌する副腎を表す振動子からなる結合2振動子系を用いた。各振動子間には相互に調整力が働いているが、副腎から脳への調整力がストレスホルモンを介したものである為、この調整力が大きくなることによりストレス入力が再現されると考える。
数理モデルを用いたシミュレーションにより、明期のストレスは末梢時計の位相を進め、暗期のストレスは位相を遅れさせるという実験結果における傾向が再現された。今後は振動子を増やしたモデルを用い、より正確に実験結果を再現できるかどうか検討する。
[1] Yu Tahara et al. (2015), Scientific Reports, 5, 11417
接触抑制を考慮した細胞増殖の粒子法による数理モデル化 平 祐介
私たちのからだは細胞が集まることで形作られている。からだや器官を構成する一つ一つの細胞は成長と分裂を繰り返すことで増殖する。細胞増殖は器官の成長や傷の治癒など私たちのからだの形成に必要なため重要な機構である。一方で、過度の細胞増殖は腫瘍や臓器の過形成といった問題を引き起こしてしまう。したがって、細胞増殖が必要以上に進行することを抑制する機構も同様に重要であるといえる。上皮組織などの細胞は一層で最密状態となるまで増殖し、以降は増殖ができない。このような細胞数増加に伴う増殖率の低下を引き起こす増殖抑制機構として接触抑制が考えられている。本研究では接触による「細胞が受ける力」に注目し、細胞が受ける力のみを考慮することで増殖抑制に関して数理モデルを用いて説明することを目的とする。
本数理モデルでは細胞を粒子と仮定し、細胞の増殖と再配置についてモデル化する。細胞は推進力を持たないと仮定しているが、細胞同士が離れている場合には近づき、細胞サイズより近づいている場合には離れるという再配置を行い、お互いが接触する距離を保とうとする。増殖率は細胞が受ける力に依存する変数によって決定する。細胞が受ける力として本研究では、細胞同士が接触することで受ける力および細胞が壁と衝突することで受ける力の 2 種類の力を考える。増殖率は細胞が受ける力の関数であるとし、細胞が受ける力が大きい場合に増殖率が低くなるように設定する。シミュレーションではシャーレ内における細胞増殖を仮定する。増殖率の式に含まれるパラメータや細胞間相互作用関数におけるパラメータを調節することで接触抑制を考慮した細胞増殖をシミュレーションすることができた。このことから細胞増殖抑制において細胞が受ける力の大きさが重要な一要因であることが示唆された。
現段階では増殖率関数の形とそのパラメータは現象が説明できるよう意図的に設定している。現在モデル設定は現在取り組んでいるが、検討中の要素は「引張力」である。細胞が受ける力を「圧縮力」と考え、圧縮力が細胞増殖を抑制するというモデルの設定およびシミュレーションを完成させた。これに対して引張力とは細胞同士が接触して、細胞表面が引っ張られることで生じる力であり、引張力の発生により細胞骨格構造が変化して増殖が促進する可能性が考えられている。今後は引張力についてもモデル設定を行い、圧縮力と引張力による細胞の増殖と抑制の両面について数理モデル化及びシミュレーションによる解析を行う。
2018年度
真正粘菌変形体における周期刺激に対する輸送管ネットワークの適応 石﨑 陽
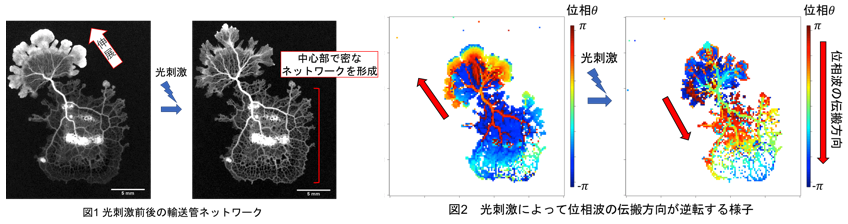
自然界に存在する多くの生物は外部からの刺激や環境変動に対して柔軟に適応する機構をもつ.単細胞生物である真正粘菌変形体(以下,粘菌と呼ぶ)もその例外ではない.粘菌はアメーバ状の生物であり,輸送管ネットワーク内を原形質が往復流動することによって栄養物質などの運搬を行う.粘菌は環境の変化や外部刺激に対して,輸送管ネットワークのパターンを変化させて柔軟に適応する.
本研究では粘菌に様々な周期の光忌避刺激を与え,その応答を解析することで粘菌の外部刺激に対する適応機構を明らかにする,その過程で粘菌の生存戦略について新たな知見が得られることを期待する.
先行研究より60分周期の光刺激下で二つの特異的な結果が得られた[1].まず,ネットワークの粗密性を評価するメッシュ度が最大化され,より密なネットワークを形成することが示唆された.さらに,刺激をきっかけに粘菌の伸展方向が高頻度で転換した.60分周期という中間的な刺激下では,密なネットワークを複数方向に伸展させることによって環境変動の頻度の高い不安定な環境における生存確率を高めようとしている可能性が示唆された.
本研究では60分周期条件でみられる特異的な応答が,光刺激による厚み振動現象の変化によって引き起こされると仮定し,振動現象における角振動数と位相に着目した解析を行う.結果から,60分周期条件では光刺激による位相波の逆転や,それに伴う伸展方向の転換がその他の周期に比べ高頻度で起こることが分かった.さらに角振動数の解析から刺激後30分以上経過すると粘菌の振動は定常状態へと遷移し,その後急激な変化は見られないことが分かった.これらの結果から,仮説として60分周期条件では位相波の逆転や伸展方向の転換によって粘菌先端部から中心部へ原形質が押し戻され,中心部でのメッシュなネットワークを保持しながら伸展する可能性が示唆された.
今後,粘菌中心部におけるメッシュ度の定量や,よりミクロな分析を行うことによって以上の仮説を検証する予定である.
[1]豊田悟史, 光刺激状況下における真正粘菌変形体のネットワーク形成, 早稲田大学先進理工学研究科, 2015年度修士論文
運動性シアノバクテリアによるコロニー形成と増殖活性の関係 溝江 恵太
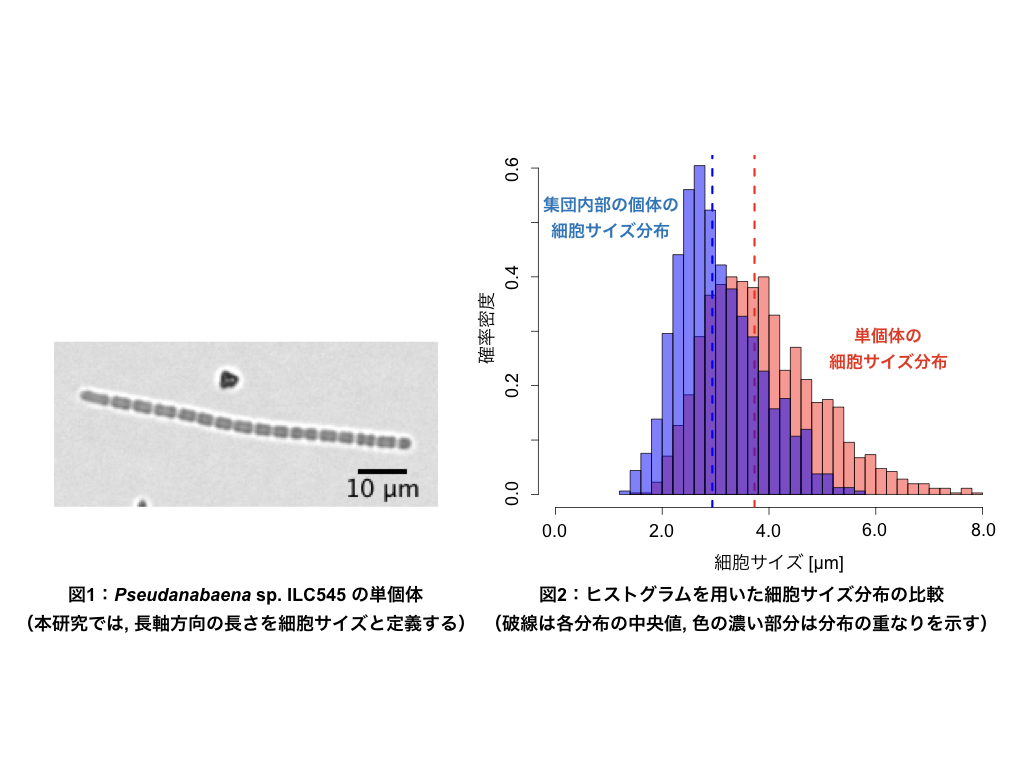
自然界において生物はしばしば集団を形成し生活している. 個体同士の間には密な相互作用が存在し, この作用による集団化で生物は防衛や生殖などの面で様々な利益を生み出している. 鳥類や魚類では集団で行動することで天敵への警戒力を上げ、アリなどの社会性昆虫では餌の探索や子育てなどの役割を分業するなど, 生物種によって集団化の目的が異なる. 本研究で扱うPseudanabaena sp. ILC545 (以下, Pseudanabaena sp.と表記)も集団を形成する生物の一種である.
本研究はPseudanabaena sp.の集団形成が細胞増殖に与える影響を明らかにすることを目的とする. Pseudanabaena sp.はムコ多糖類を成分とする液状物質(以下, 粘液と表記)を分泌することが分かっており, 先行研究によって粘液が自身の増殖率を上昇させる可能性が示唆されている. 個体数が多い集団では粘液の存在量が増加し, 分裂が活発になることで小さいサイズの細胞が多く観測されるのではないかと考え, 個体の細胞サイズに着目し指標とする.
単個体で滑走するPseudanabaena sp.および集団内部に存在するPseudanabaena sp.の細胞サイズ分布の比較より, 単個体で滑走する場合と集団形態をとった場合では観測される分布に差異が見られた. この結果から集団形成がPseudanabaena sp.の細胞増殖に影響を与えていることが示唆されたが, 粘液が直接的に影響しているかは明らかでない. 現在, 追実験および簡単なシミュレーションを用いることで増殖に影響を与える機構の解明に取り組んでいる.
自動運転車の反応時間を考慮した高速道路における渋滞のシミュレーション 戸田 賢
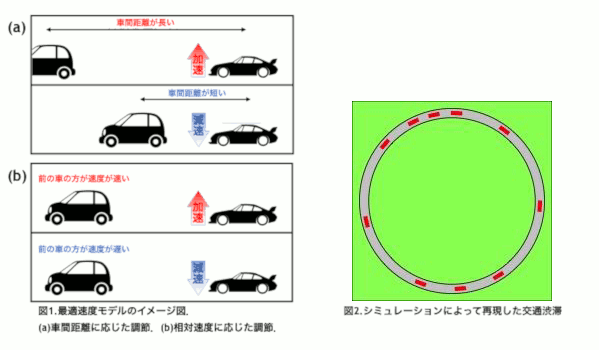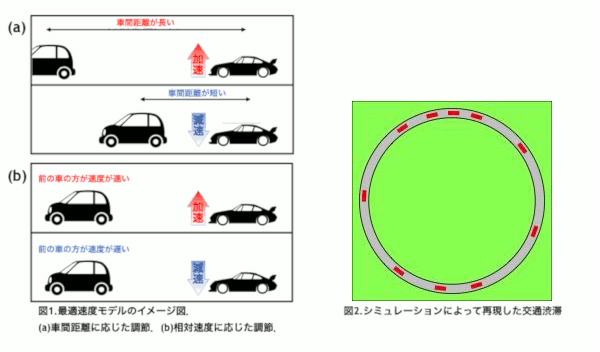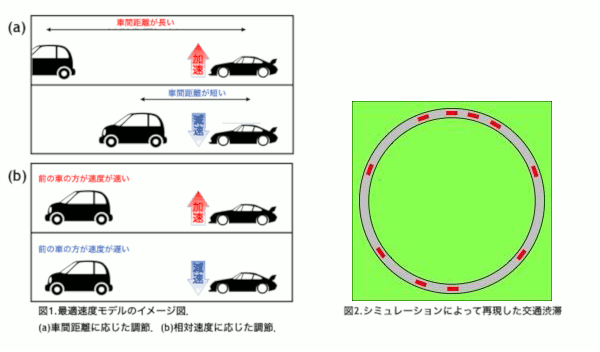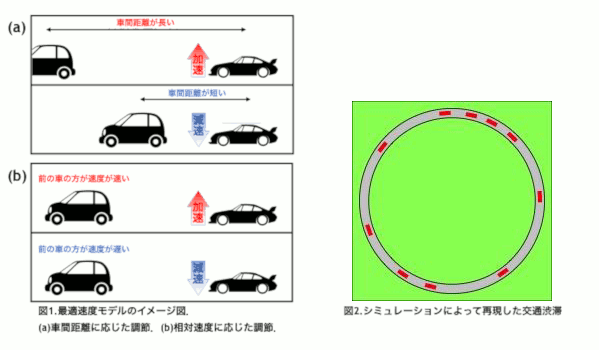
東京オリンピックを迎える2020年を目処に,革新的な交通技術である自動運転車の導入が予定されている.自動運転車はドライバーが運転に関与することなく運転操作を制御し自動で走行することを目指す乗用車である.事故率の低下や交通渋滞の軽減が期待されている反面,交通が乱れることが懸念されている.本研究では高速道路の渋滞に着目し,自動運転車の普及によって渋滞が改善されるのか,流れにどのような変化が表れるのかをシミュレーションによって解き明かすことを目的とする.
普通車と自動運転車を区別するために,反応時間に着目した.反応時間とは認知から行動までにかかる時間のことで,人間の運転する普通車よりも自動運転車の方が短いことが知られている[1].本研究では交通流を再現する最適速度モデル[2]に時間遅れを導入し,反応時間の違いを組み込むことで普通車と自動運転車を区別した.
自動運転車の割合を変化させたシミュレーションを行った結果,自動運転車が10〜70%の範囲で0%の場合よりも加減速の頻度が高くなるパラメータの存在が確認された.このことから,自動運転車が導入された初期段階において普通車のドライバーへの負担が増える可能性が示唆された[3].
以上の結果を踏まえて今後の展望としては,より現実的な車の加減速性能を元としたモデルの再検討や,車線変更を再現したシミュレーション等を行い,その結果を元に考察を行いたいと考えている.
[1] Schakel, W. J., Van Arem, B., & Netten, B. D. (2010), Intelligent Transportation Systems (ITSC) , 13 759-764.
[2] Bando, M., Hasebe, K., Nakayama, A., Shi- bata, A., and Sugiyama, Y. (1994), Japan Journal of Industrial and Applied Mathemat- ics, 11(2), 203-223.
[3] 戸田賢, 高松敦子, ”自動運転車と人間が運転する自動車の混在下で発生する渋滞シミュレーション”, 第23回交通流と自己駆動粒子系のシンポジウム, 23, 79-82, 2017.
運動性シアノバクテリアの往復運動の解析 幸﨑 峻
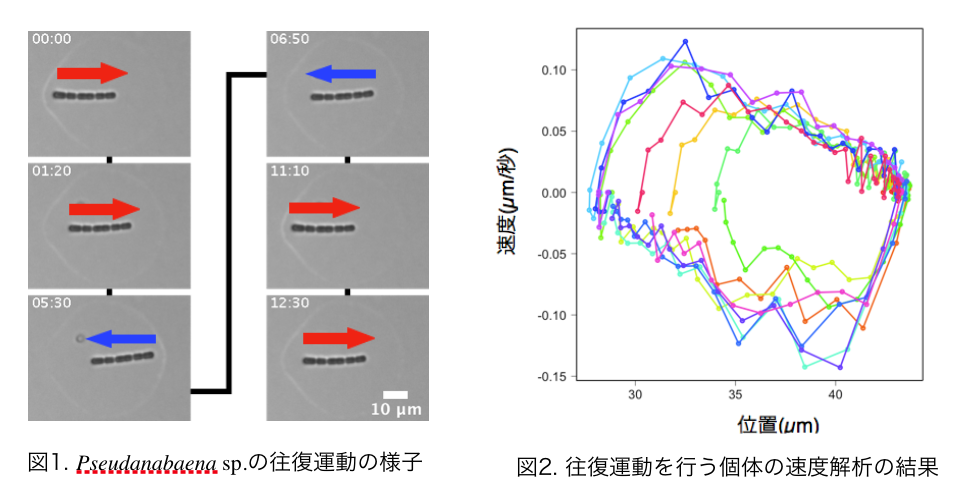
研究対象であるPseudanabaena sp. ILC545株は, 複数の細胞が連なって個体を形成する運動性の鎖状シアノバクテリアである. 単独では培地上を滑走運動し, 時折進行方向を反転する. 複数の個体が集まると特徴的なコロニーパターンを形成する. しかし, 詳細な運動機構は未知である. Pseudanabaena sp.のコロニー形成のメカニズムについて考察するために, 運動機構を明らかにすることを目指している.
Pseudanabaena sp.は運動に伴い, 液状の粘性物質(以下, 粘液と呼ぶ)を放出する. 培養されたバクテリア集団の周りには多量の粘液が存在し, 集団を離れた個体の周りには少量の粘液がカプセル状に存在する. 短い個体は粘液カプセルの外部へ進出ができない. 従ってこの粘液カプセルは個体が移動可能な閉じた領域となる. この内部で観察されるPseudanabaena sp.の特徴的な往復運動について詳細な解析を行っている.
粘液カプセルの両端(以降, 粘液の境界と呼ぶ)で個体が進行方向を反転した後, 運動速度が急峻に立ち上がる. 粘液の境界がバネのような役割を果たしていると考え, 以下のような個体の運動を想定する. 方向転換直後に粘液の境界からバネの力を受け, 個体の速度は最大となる. 個体が粘液の境界から離れることで, 個体の速度は駆動力による一定の速度に落ち着く. それから反対側の粘液の境界にぶつかることで, 再びバネを伸ばしながら個体の速度は低下する. 最終的に個体は停止し, 方向転換を起こす. この運動が繰り返され, 往復運動が観察されている. このような想定を元に運動方程式を仮定し, 駆動力等のパラメータ推定の結果からPseudanabaena sp.の運動機構に知見を得ることを期待している. 現在までの研究でPseudanabaena sp.は双方向性の運動機構を持ち, その進行方向に依らず個体の駆動力が一定であることが示唆された.
真正粘菌変形体の二者択一問題における行動決定パターンの再現 満武 雄也
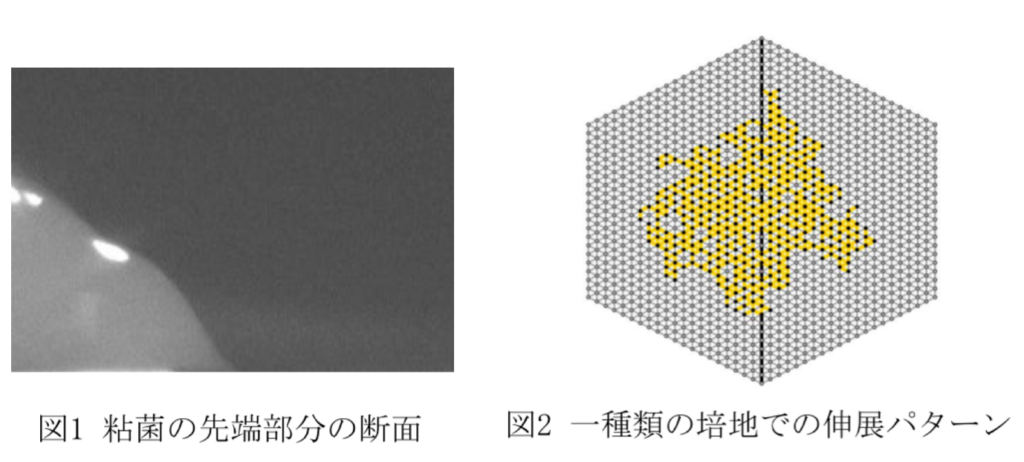
真正粘菌変形体(以下,粘菌と略す)は,一細胞の大きさが10µm〜1m程の多核の単細胞生物である.粘菌は、高度に発達した脳や神経系を持たないが,複雑な環境下でも行動することができたり,迷路を解くことができたりするなどまるで知性があるかのように振る舞う.
本研究では,粘菌の複雑な環境下においても行動できる仕組みについて知見を得ることを目的とする.そこで,粘菌に二者択一問題を与える.具体的に,シャーレを異なる二種類の環境培地で分割し,その境界で粘菌の伸展を観察する.特に,粘菌が片方の培地を選択して伸展し始めるまで(=行動決定前)と選択した後(=行動決定後)の伸展パターンの違いについて,粘菌の先端部分と培地のなす角度(=接触角)に着目してアプローチする.
実験の結果,粘菌は最終的に接触角が小さい方向へ成長し,行動決定前に比べて行動決定後の方が接触角は小さくなることが分かった.これは,一方の培地へ伸展できることにより細胞内の原形質圧が低下したためと考えられる.さらに,二種類の培地における接触角の差が小さいほど,行動決定前期間が長くなることも分かった.
以上の結果を元に,ネットワーク成長モデル[1]を参考に数理モデルを構築し,二者択一問題における伸展パターンの再現を目指す.管の伸展の指向性と伸展確率を表すパラメータを用いたルールに加えて,行動決定前後における伸展のしやすさの違いを考慮する.その際,行動決定前期間は左右の接触角の差に依存して変更する.さらに,粘菌には体積を一定に保ち伸展するという特徴がある[2].その為,伸展のみならず管の衰退も今後は考えていく.
[1] Takamatsu, A., Takaba, E. and Takizawa, G. (2009) “Environment-dependent morphology in plasmodium of true slime mold Physarum polycephalum and network growth model”, J. Theor. Biol., 256, 29-44.
[2] Takamatsu, A., et al (2017) “Energy-saving with low dimensional network in Physarum plasmodium”, J. Phys. D: Appl. Phys., 50, 154003
2019年度
微小粘菌における空間探索の環境依存 稲田 英幸
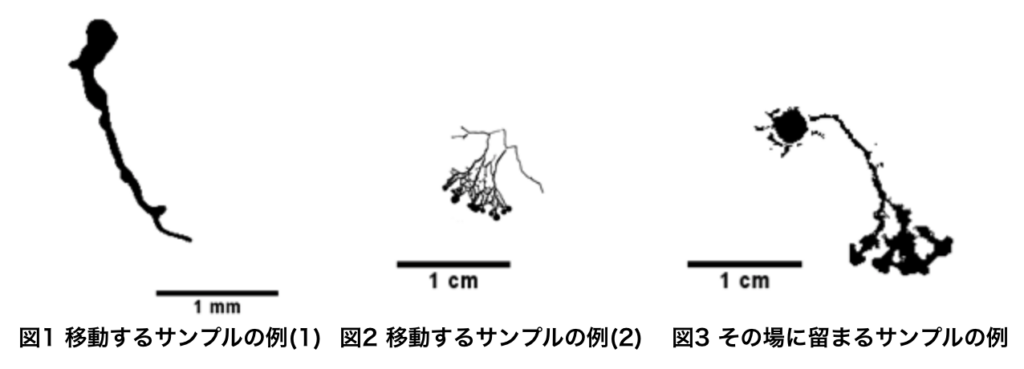
本研究で用いた真正粘菌変形体(以下, 粘菌と略す)は, 多核の単細胞生物である. 粘菌は管状のネットワークを形成し, 原形質流動によって栄養の運搬を行っている. このネットワーク構造は, オートミール等粘菌が好む物質を含む環境ではメッシュ状, KCl等粘菌にとって好ましくない物質を含む環境では樹状と, 周りの環境に応じた形態をとることが知られている.
先行研究から, このネットワーク構造は粘菌の大きさが影響していることが示唆されている. [1]そこで本研究では小さな粘菌に着目し, 大きさが粘菌の形態にどのような影響を与えているのかを解析し, 粘菌がなぜ戦略的な空間探索を行うことができるのかを解明することを本研究の目的とした.
本研究では, 粘菌の管を切断した際に流れ出る原形質をマイクロピペットを用いて吸い取ることにより, 体積の異なる粘菌を調整し実験を行った. その結果, いずれの体積においても実験開始直後は円形を保ったまま成長することは共通していた。しかしその後の探索の仕方については体積によって違いが見られた1µl~3µl程の粘菌では、実験開始時に粘菌を置いた位置から中心座標を変え, 移動するような探索を行ったのに対し, 体積が増加していくと中心座標を変えないまま, 軸索のようなものを伸ばし探索を行っていた.このことから粘菌が自らの大きさを把握し, 大きさよって空間探索の方法を変えるということが示唆された.
今後はさらに小さい粘菌で同様の実験を行うこと, 培地条件を通常培地から誘引培地や忌避培地に変える, 温度や湿度を変える, など様々な環境条件で実験を行うことによって, 微小な粘菌における空間探索の環境依存について知見を得ることを目標としている.
[1] Takamatsu, A., Takaba, E. and Takizawa, G. (2009) “Environment-dependent morphology in plasmodium of true slime mold Physarum polycephalum and network growth model”, J. Theor. Biol., 256, 29-44.
マウス概日時計における細胞の個性を考慮した数理モデルの構築 岩崎 真依

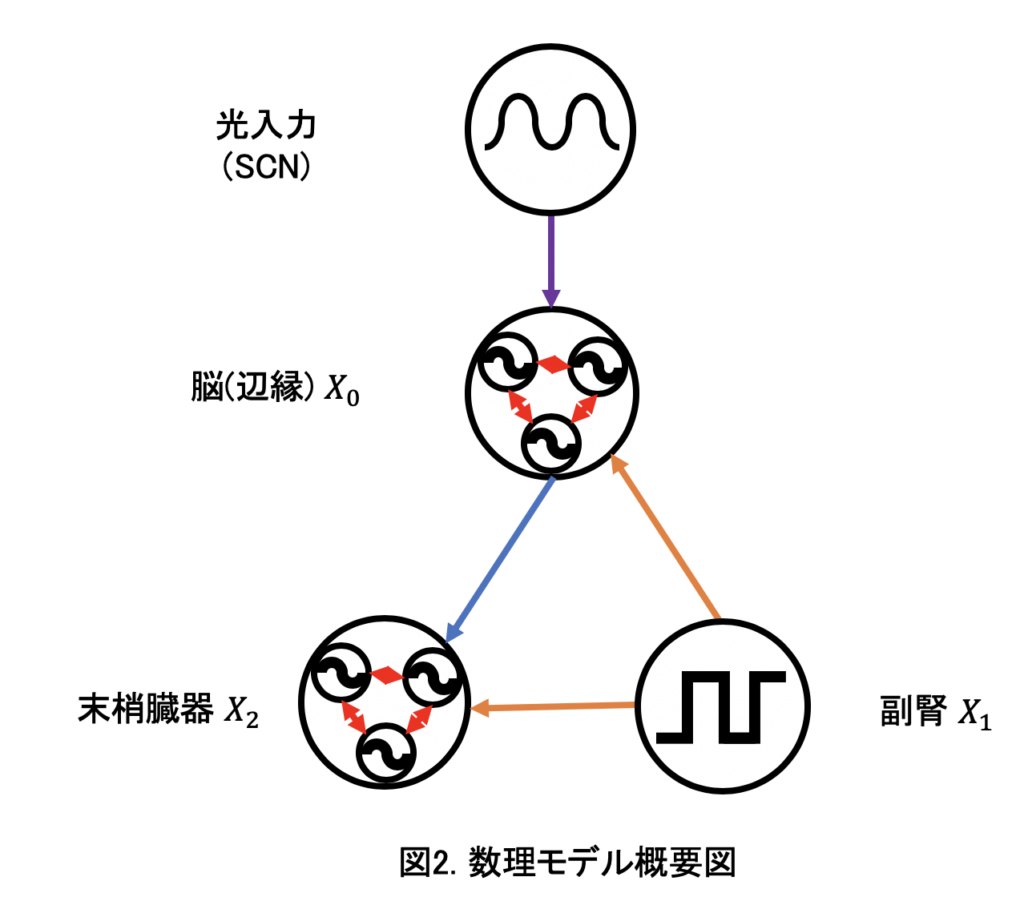
生物には約24時間のリズムを刻む概日リズムという機構がある. このリズムは身体的・精神的ストレスによって大きく乱れる. 特に,特異的な時刻におけるストレスは,末梢臓器(腎臓)の概日リズムを消失させてしまうことが知られている[1]. このリズムが消失する原因としては,ストレスによって細胞のリズムが乱れること(脱同調)が原因であると考えられているが[2],細胞単位の概日リズムに着目したストレス負荷実験はなされていない. そこで本研究では,細胞単位の数理モデルを構築し,細胞の概日リズムがストレスによって乱れることによって,臓器の概日リズムが消失することを再現し,そのメカニズムを探る.
本研究では脳(辺縁)と末梢臓器で構成される数理モデルを構築する.各臓器はそれぞれリズムを持つ細胞集団で構成される.末梢臓器の細胞集団にはストレスホルモンが作用し,脳の細胞集団にはそれに加えてSCNを経由した光刺激が入力される.更に,臓器内の細胞同士は相互作用する(図2).
本モデルで解析した結果,ストレス応答の感受性を適切な値にすることによって特定時刻のストレス刺激により末梢臓器の細胞が脱同調する様子が再現できた.つまり,脳とストレスホルモンを出す副腎からの入力のバランスによって,末梢臓器(腎臓)でリズムが消失したと考えられる.また,リズム消失が見られない臓器(肝臓・顎下腺)においてもストレス応答の感受性の値を変化させることによって、実験結果を再現できた.
今後,ストレスホルモンを分泌する副腎の日内リズムを考えて,より現実系に近づけていく予定である.
[1] Yu Tahara et al. (2015), Scientific Reports, 5, 11417
[2] Hideaki Ukai et al. (2007), nature cell biology, 9, 1327-1334
真正粘菌変形体における輸送管ネットワークの流動解析 稲井 由美

本研究で用いた真正粘菌変形体(以下, 粘菌と略す)は, 輸送管ネットワークを形成し, 管内を原形質が往復流動する. 管は原形質の流量が多いほど太くなり, 少ないほど細くなる性質をもつ. この性質は管の成長方程式として数理モデル化されている. モデル式には粘菌の管の成長過程を特徴づける4つのパラメータが含まれており, それらの値の違いは成長過程の違いを表す. 本研究ではこのパラメータ値を見積もり, 管の成長過程を定量的に評価すると共に, 培地環境の違いによる粘菌の成長の差異を明らかにすることを目的とする.
実験では, 図1のような長さの異なる二本の管を形成させる系と, 図2のような自由なネットワークを形成させる系のそれぞれにおいて管の半径と原形質の流量を測定し, それらの値を用いてモデル式に含まれるパラメータの推定をした. 図1の系において, 管の両端にかかる圧力差と全流量の時間変化を考慮した場合と, 考慮しない場合の二通りの方法でパラメータの推定をしたところ, どちらも同程度の値を得ることができた. このことから, パラメータの推定において圧力差や全流量の時間変化は影響しないことが示唆された. そこで, 図2の系においても圧力差や全流量の変化を考慮せずにパラメータの推定をした.
今後は図2の系において, 培地環境を変化させ, パラメータ値の違いから, 原形質の流量に対する管の太さの成長過程の違いを明らかにし, 粘菌の環境適応を定量的に評価する.
クロヤマアリのエサ運搬時における行動変化 和田 優也

アリは採餌行動を起こすことが知られていて,その際にエサの運搬方法が変わることが知られている.その運搬方法について調べるために実験を行ったが、その実験対象にクロヤマアリを用いた.クロヤマアリは日本全国に分布するアリでとても馴染み深いアリである.クロヤマアリ(以下アリと略す)においては二種類の単独運搬と協同運搬が実験で見られている.エサを単独で前向きに持ち上げながら運ぶ「前向き運搬」,エサを単独で後ろ向きに引きずりながら運ぶ「後ろ向き運搬」,複数個体で引きずりながら運ぶ「協同運搬」の三種類である.本研究においては,これらの運搬方法がどのような条件で切り替えられるか,それぞれの運搬方法にはどういった差異があるかどうかについて言及することを目的がである.
実験方法は,実験場にエサ(今回は桜エビを使用)を置き,アリがエサを運搬している様子をカメラで撮影してデータの解析を行った.実験結果からエサの重さによる切り替えが一番の要因であると考えた.しかし,前向き運搬と後ろ向き運搬ではある一定の重さで切り替えが起こったが,協同運搬においてはどの重さでも常に確認出来た.つまり,単独運搬は切り替えが起こるものの協同運搬においては常に発生の可能性があり,軽いエサにおいても常に運搬に加わろうという性質があることが分かった.他にも,単独運搬においては,前向き運搬と後ろ向き運搬で運搬速度が変わることが分かり,前向き運搬の方が運搬速度が速い結果になった.
今後,障害物に対する反応性やエサの形による反応性など様々な指標を用いて解析を行い,新たな特徴量を見つけ出したい.
2020年度
真正粘菌変形体における馴化の要因 米岡 笑里
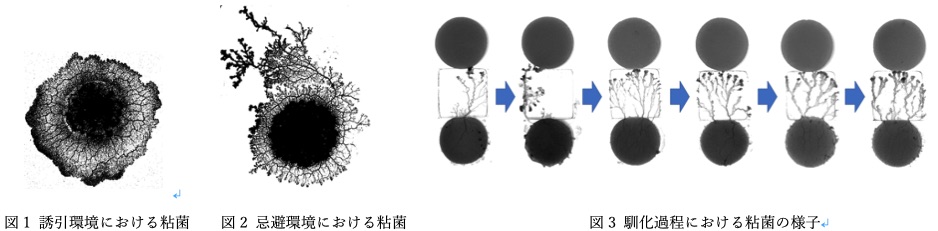
生物が誕生してから環境は目まぐるしく変化し,その環境変化に適応することのできたものが生き残ってきたと考えられている.つまり現存している生物は,厳しい環境を含めた様々な環境に適応することのできる有利な性質を持っているとされている.本研究では,どのようなメカニズムにより生物が厳しい環境に適応していくのかについて,真正粘菌変形体(以下,粘菌と略称する)を対象として研究を行う.
粘菌は多核の単細胞生物であり,輸送管ネットワークを構築する.このネットワーク形態は環境に応じて変化する.また先行研究から,粘菌は忌避物質に対し順化を示すことが示唆された[1].さらに,粘菌における順化の要因は輸送管ネットワークにあることが明らかになった.本研究では,順化過程における粘菌のネットワーク形態を観察し,順化のメカニズムを明らかにすることを目的とする.具体的には,数日間連続で粘菌に忌避物質を与えることで順化をさせ,その過程において管の配置がどのように変化するかを解析した.
実験の結果,順化過程で伸展方向に並行な太い管が現れることによって粘菌は順化を示すことが示唆された.
[1] Boisseau,RP.,Vogel,D.andDussutour,A.(2016)”Habituationinnon-neuralorganisms: evidence from slime moulds”, Proc. R. Soc. B,283.
運動性シアノバクテリアにおける集団パターン形成のモデル化 大友 和理
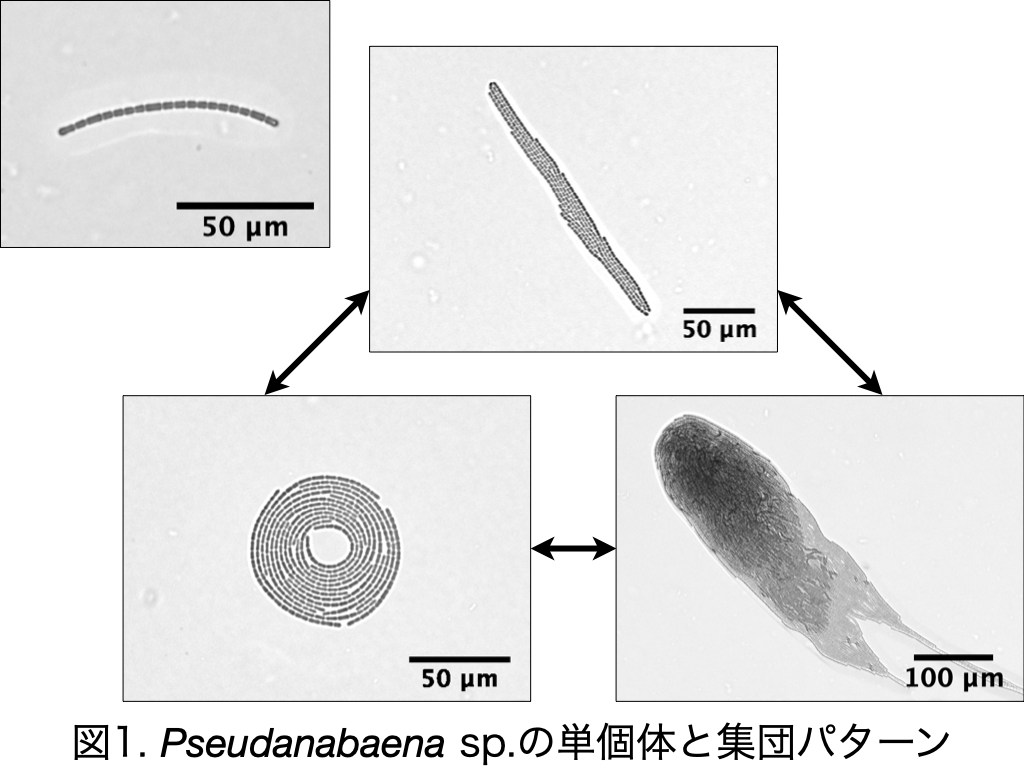
自然界において生物は秩序的な集団パターンを形成することがある.集団の形成は哺乳類や魚類だけでなく,バクテリアでも見られる.バクテリア は集団を形成することで効率よく栄養を獲得したり,胞子となって休眠状態になったりすることができる[1].バクテリアにとって集団を形成することは種の生存に有利に働くと考えられる.
本研究で扱う運動性シアノバクテリアのPseudanabaena sp. ILC545 (以下,Pseudanabaena sp.と表記) も,培地上を滑走運動しながら集団化することで三種類の集団パターンを形成する(図1)[2].それぞれの集団パターンは自発的に崩壊したり,集団同士が衝突したりすることで各形態間を流動的に遷移する.こうした集団パターンの形成や遷移に関わる本質的なメカニズムは明らかとなっていない.そこで,本研究ではPseudanabaena sp.が多様な集団パターンを形成するメカニズムを明らかにすることを目的とする.
本研究では鎖状に連結させた自己駆動粒子でバクテリア単個体を数理モデル化する,数理モデルでは次に述べるような,Pseudanabaena sp.に見られる特徴的な現象を考慮する:1. 駆動力は個体の長軸方向に維持されやすい.2. 個体間には引き合う相互作用が働く,3. 個体が分泌する粘液によって抗力が低減され速度が上昇する.シミュレーション空間上でモデル化した個体を多数運動させることで数値実験を行う(図2).
現在までに,パラメータを変化させることで多個体が集団化していく様子も変化することが確認された.今後は集団の大きさや配向度合いを指標として,各パラメータが集団パターンに与える影響を定量的に解析する予定である.
[1] 不藤亮介 (2013) “粘液細菌; この可憐で賢き狩人たち”, 生物工学, 91, 532–353.
[2] 高松敦子, 工藤亮太, 齋藤拓, 木村拓 (2014) “運動性シアノバクテリア集団の遷移現象”, 第 20 回交通流シミュレーションシンポジウム論文集, 23–26.
真正粘菌変形体における輸送管ネットワークの分岐則 柴田 将広


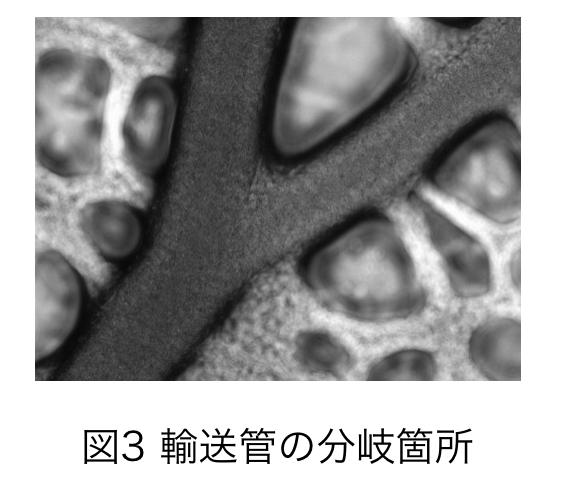
物質やシグナルを伝達する輸送管ネットワークは、血管や葉脈などあらゆる生物で見られる.生物で見られる輸送管ネットワークにおける管の分岐則としてマレー則がある.マレーは,管の消費エネルギーが最小化されるとき,管内の流量が半径の3乗に比例し,その結果,管径は3乗則に従い分岐することを示した[1].
本研究の対象である真正粘菌変形体(以下,粘菌と略す)も輸送管ネットワークを構築する.粘菌は大きさが10µm〜1m程の単細胞生物である.粘菌の特徴の一つに,環境に応じて輸送管ネットワーク形態が変化することがある.本研究では,様々な環境の粘菌においてマレー則が成り立つか調べ,管の分岐則について知見を得るとこを目的とする.具体的には,粘菌を様々な環境で培養し,分岐において管内の原形質流動の様子を撮影した.この動画から流量,半径を算出し,両者の関係を推定した.
実験の結果,環境によらず,粘菌が進展方向を決めた後は,管内の流量が半径の3乗に比例しマレー則が成り立つことが,進展方向を決める前は,3より低い指数を持つことが明らかとなった.
[1] C.D.Murray(1926), The physiological principle of minimum work I, the vascular system and the cost of blood volume,Proc. Natl.Acad. Sci. USA 12,207-214.
一次元状の真正粘菌変形体における時空間振動パターンの解析 三木 渚

地球上の生物を取り巻く環境は時事刻々と変化している。生物はこの様な環境変化が起きる度に進化しつづけることで現在まで生存してきた。生物の環境への適応性は生命の維持に必要不可欠なものである。 多核の単細胞生物である真正粘菌変形体(以下、「粘菌」と呼称する)はゲル状の輸送管と原形質ゾルから構成されるアメーバ状細胞であり、大きいもので1mほどの大きさになる。輸送管内を原形質が往復流動することで栄養物が運搬され、この往復流動によって粘菌は周期的に振動する。加えて、粘菌は環境への適応性を容易に可視化できる特徴を持つ。粘菌の輸送管ネットワークは環境に依存して様々な形態をとる。成長に伴ったネットワーク形成時に、厚み振動の時空間パターンと振動数が変化する事が明らかとなっている。しかしながら、これらの変化がネットワーク形状の変化によるものなのかネットワークサイズによるものなのか、明確でない。本研究ではこれらの振動変化がおこる仕組みを明らかにすることを目的とし、ネットワーク形状を1次元状のリングとして単純化し、時空間振動パターンのリングサイズ依存性を調べる。 具体的には図のように様々なサイズの1次元リング状の粘菌を形成して粘菌の時空間振動パターンを解析する。現在までに全てのリングサイズの粘菌において回転振動が観察され、リングサイズに依存して振動パターンに変化が生じることを確認した。 今後は粘菌の振動パターンがリングサイズにどのように依存しているのか、より定量的に解析を進める所存である。
孤立した上皮細胞の運動と形態の解析 柴垣 志文
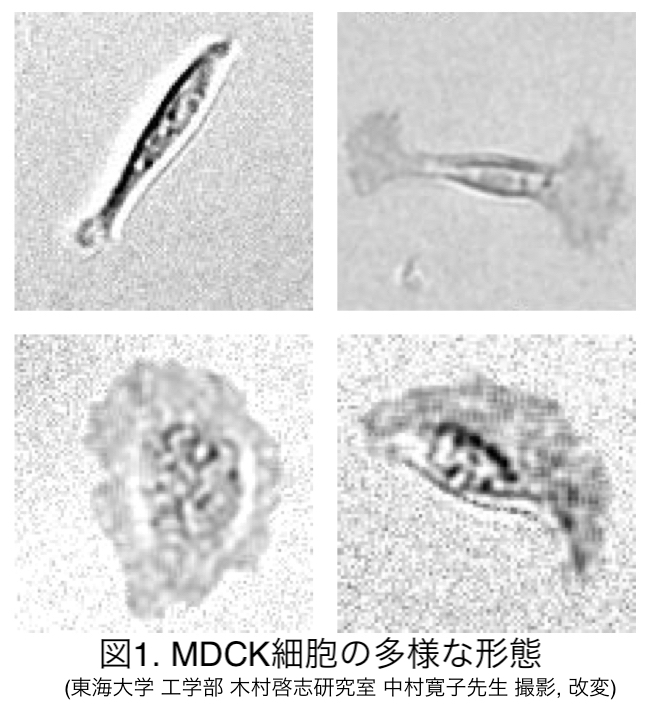
生体内の細胞は基本的に集団となって機能する。一方で集団を構成している個々の細胞が持つ性質については見落とされがちである。 単体で存在する細胞は細胞種によって多様な運動や形態を示すことが複数の細胞種については知られている[1,2]。さらに個々の細胞の有する動態は組織形成や傷の修復において重要な役割を果たすことが示されている[3,4]。 本研究ではイヌ腎臓尿細管上皮由来の細胞(Madin-Darby canine kidney; MDCK cells)を用いた。なお培養・実験データの提供は共同研究者の中村らのご厚意によって行われている。本研究では現在までにMDCK細胞において、同一細胞種でありながら多様な運動・形態を示すことが見出されている。そこで同一細胞種から発生する多様な動態は、細胞内の同一の機構によって生成可能であるとの仮説を立て、本研究では単体の細胞の動態を定量的に把握することを目的とした解析を行っている。
実験観察から同一培養条件下の同一細胞種ながら多様な動態パターンが観察された(図1)。また木村らの実施したクローニング実験(細胞集団内の遺伝情報をそろえる実験)では、由来遺伝子によって観察される動態パターンに顕著な差は認められなかった。さらに動態パターン間を自発的に遷移する現象が観察されているため、多様な動態パターンが遺伝子のみでは説明できず、細胞内外における物理的な相互作用を考慮した説明が必要である。現在、当研究室の三瀬らにより一細胞の動態を表現する数理モデル構築および解析が行われている。また当研究室の八塚らによる機械学習を用いた細胞形態の自動分類が行われ, 定性的であった細胞形態の分類を客観的・定量的に評価することを目指している。共同研究者の松永らのご好意により教授していただいた細胞培養・実験方法の指導に基づき、当研究室においても培養・実験を行える環境を構築した。現在は当研究室の岡本らにより同様の細胞を用いて培養条件を変化させた際の細胞動態に関する解析が行われている。
将来的には現在用いている細胞種以外の細胞についても動態パラメータを定量・同定することで細胞の有する性質を動態から推定し、実験に必要な試薬量の最適化や病理診断などへの応用が期待される。 (共同研究先:東海大学 工学部 木村啓志研究室 中村寛子先生(培養動画の撮影・提供), 東京大学 生産技術研究所 松永行子研究室 中島忠章先生(実験・培養方法の指導))
接着斑とストレスファイバーを考慮した細胞の運動と形態のシミュレーション 三瀬 翔太
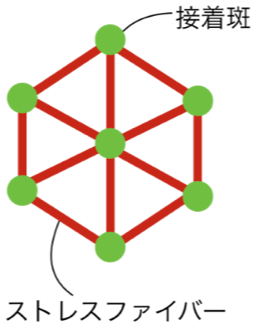
細胞は基質の上で様々な運動パターンを示す。例えば、マウス線維芽細胞では往復運動が、魚類表皮由来のケラトサイトでは直進運動が、細胞性粘菌では停滞状態から直進運動への変化が報告されている[1,2,3]。当研究室の柴垣らは、イヌ腎臓尿細管上皮細胞由来のMDCK細胞(Madin-Darby canine kidney cell)が同一培養条件下でも上記と同様の運動パターンがあり更にパターン間を遷移することを見出した(この研究の詳細については柴垣がポスター発表を行う)。従って、これらの複数の運動パターンが同一の運動機構でも生成できる可能性がある。細胞と基質を接着させる接着斑とそれらを接続するストレスファイバーによって細胞は駆動される[4]。そこで、本研究では図のような接着斑とストレスファイバーで一細胞を表したシンプルな数理モデルを構築した。 本モデルでは、接着斑を質点、ストレスファイバーを質点間のバネと仮定した。質点には接着斑を形成する接着構造の数に応じて細胞の外側へ伸展する力を与えた。接着構造の数はバネの張力に応じて増減させた。 解析の結果、細胞の停滞状態から直進運動への遷移が見られた。 今後は数理モデルを改良し、その他の運動パターンの遷移条件を探る予定である。発表では本モデルと解析結果の現状について、現実の細胞と比較しながら議論する。
[1] Ebata, H., et al. Sci Rep, 8, 5153, (2018).
[2] Keren, K., et al. Nature, 453, 475. (2008).
[3] Artemenko, Y., et al. CMLS, 71, 3711-3747, (2014).
[4] Burridge, K., & Guilluy, C. Exp Cell Res, 343, 14-20. (2016).
2021年度
照射光の照度と波長がシアノバクテリアの運動と形態に与える影響 矢口 風香
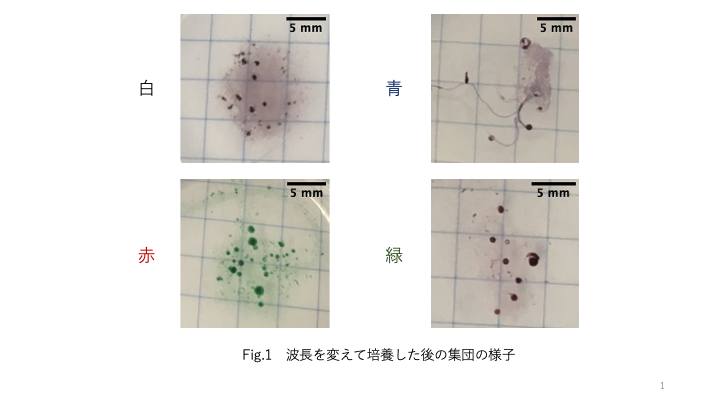
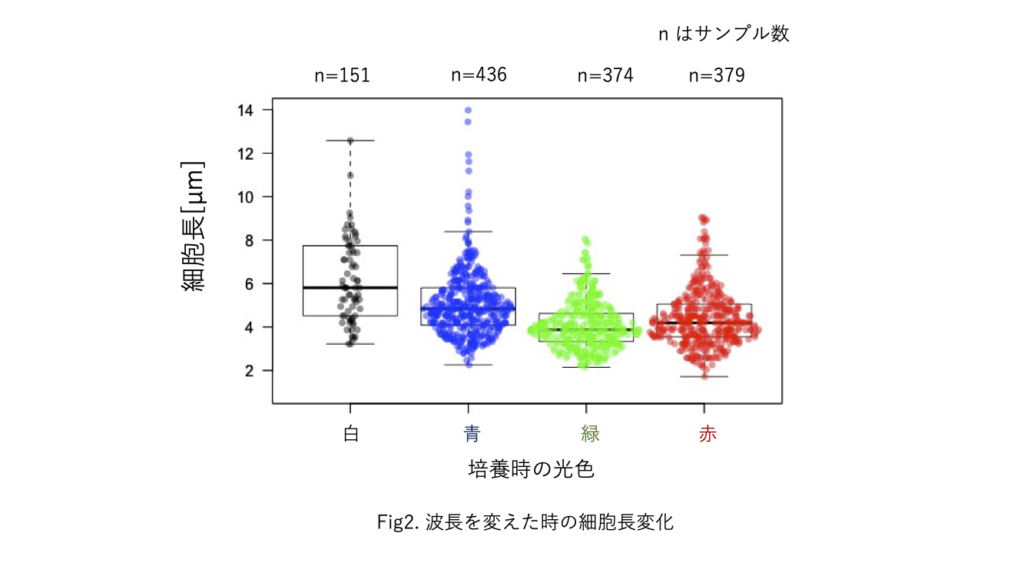
太古より続く激しい生存競争に勝ち抜くために, 生物は特徴的な性質を備える. 環境に適応できるかどうかはバクテリアから哺乳類まで生存に関わる重要なことである. 本研究ではバクテリアの環境適応を観察している.
私の研究対象である運動性シアノバクテリアPseudanabaena sp. NIES-4403(以下Pseudanabaena sp. と表記)は原核光合成生物である. Pseudanabaena sp. には補色馴化と呼ばれる環境の光に適応する性質が備わっていることが知られている[1]. そこで, 本研究では光環境が変化した時にPseudanabaena sp. 単個体に生じる変化に着目する. Pseudanabaena sp. を異なる明るさや波長の照射光下で培養し, 単個体の運動速度や個体長、細胞長を計測する. それらのデータを統計的に分析することで, Pseudanabaena sp. の運動や形態に及ぼされる影響を調べる.
解析の結果から, 光の波長によって細胞長が影響を受けている傾向が示された. 観察の結果から, 波長を変えて培養した時の集団の様子にも影響があることがわかった. 今後はPseudanabaena sp. 特有の3種類のコロニー形態を比較し, 光環境がPseudanabaena sp. に与える影響を多個体で観察していく.
[1] 深澤有貴(2012) “運動性シアノバクテリアのコロニーパターン形成のダイナミクス”, 早稲田大学大学院 先進理工学研究科 修士論文.
細胞競合による多能性維持機構の数理モデル 運動性の効果 田野口佳奈


多細胞生物は組織化された細胞集団によって形成
されているが,個々の細胞の挙動や遺伝子発現には
ゆらぎやばらつきが存在する.それにも関わらず,
正常な組織形成や恒常性維持できるのは,不要な細
胞を排除するメカニズムが存在するためである.そ
の一つに「細胞競合」がある.細胞競合は,状態の
異なる二種の細胞が隣接した状況において引き起こ
される細胞が排除される現象である.Morata と
Ripoll によってショウジョウバエの翅原基におい
て初めて発見されたこの現象は,現在ではさまざま
な状況あるいは生物種で起きることが確認されてい
る.その中で本研究では,マウス初期胚において,
Myc発現レベルの違いによって引き起こされる細胞
競合に着目する.Mycは細胞の代謝と増殖を調節す
る働きなどを持つ.マウス初期胚では,Mycレベル
が高い(Myc-H)細胞と,低い(Myc-L)細胞が一
定時間以上隣接すると,Myc-L細胞が排除される.
一方で,Myc-L細胞は多能性が低い細胞であること
が分かっているため,細胞競合により組織の多能性
を維持しているのではないかと考えられている.
マウスの初期胚では細胞が活発に運動する様子が
観察されている.この系では,細胞の運動性が高い
ほど,Myc-H細胞が多くのMyc-L細胞と遭遇すると
考えられる.一方で,高い運動性は一組のMyc-Hと
Myc-L細胞が隣接状態を維持する時間を低下させる
はずである.従って,運動性はマウス初期胚におけ
る細胞競合において,相反する二つの効果を持つ.
そこで,本研究は細胞の運動性が細胞競合に及ぼす
影響について,粒子法というシミュレーション技法
により調べることを目的とした.
真正粘菌変形体における輸送管ネットワーク形成の数理モデル 前田 航祐
ネットワーク構造は私たちの身の回りで多く見られる. 交通網や人間関係, コンピュータネットワークなどが例として挙げられ, その構造を最適化することは重要な問題である.
本研究で扱っている真正粘菌変形体(以下, 粘菌)も輸送管を張り巡らせたネットワーク構造を形成する. 粘菌は大きさが10μm-1m程の単細胞生物である. 粘菌の特徴として, そのネットワーク形態は環境によって変化することが知られている. 栄養分を多く含む培地では網目状のネットワークを形成し, 有害な化学物質を含む培地では樹状のネットワークを形成する. 網目状のネットワーク形態は栄養分の吸収効率を高め, 樹状のネットワークはより良い環境を探すのに役立つと考えられる.
本研究では単細胞生物である粘菌が形態を最適化するメカニズムを知るため, 数理モデルによって輸送管ネットワーク形成の再現を試みる. 輸送管の伸展や管径の変化を取り入れた数理モデルを用いることで, 粘菌同様に環境に応じて形態が変化するネットワーク形成を再現した. 細胞内の化学物質がネットワーク形成に関係していると考えられるため, 今後は数理モデルに化学物質に対応する要素の導入を考えていく.
粒子-ファイバーモデルを用いた細胞-基質間の接着性による形態制御 八塚 剛志


細胞の足場環境は, 細胞の形態や運動だけでなく, 細胞分化さえも制御することが知られている. ところが上皮細胞であるMDCK(Madin-Darby canine kidney)細胞では, 足場環境が固定されているにもかかわらず, 孤立状態にある細胞が様々な形態と運動を示す. この多様な形態と運動性を再現するモデルとして, 粒子-ファイバーモデル(図1)が提案された[1]. 本研究では, 足場環境(細胞-基質間の接着性)が細胞形態を制御するメカニズムを明らかにするため, 実験結果と比較しながら, 粒子-ファイバーモデルを用いた解析を行う.
モデル解析に先立ち, パラメータ探索の基準を定めるため, 先行研究で取得した培地環境固定条件における, MDCK細胞の定量的な形態解析を行った. ここでは, 物体検出アルゴリズムYOLO(you only look once)[2]を適用した. 本手法により, 細胞の位置検出と形態分類を, 客観的・自動的, かつ高速に行うことが可能になった(図2).
次に, 数理モデルを用いて, 接着・脱離確率を制御するパラメータを段階的に変化させた場合の応答を解析した. その結果, 形態は接着確率よりも脱離確率に強く依存して変化する傾向を示した. 以上より, 細胞-基質間の接着性は細胞の形態と遷移現象に関する重要な制御パラメータであることが, 粒子-ファイバーモデルを用いて確かめられた.
今後は本モデルを用いて, 接着性による形態制御のメカニズムについて, 更に詳細解析する予定である.
[1] S. Mise, et al., (2020) Diversity in self-organized forms and migration modes in isolated epithelial cells, Artifical Life and Robotics,25, 523-528.
[2] J. Redmon, et al., (2016) You only look once: Unified, Real-Time Object Detection, arXiv:1506.02640.
2022年度
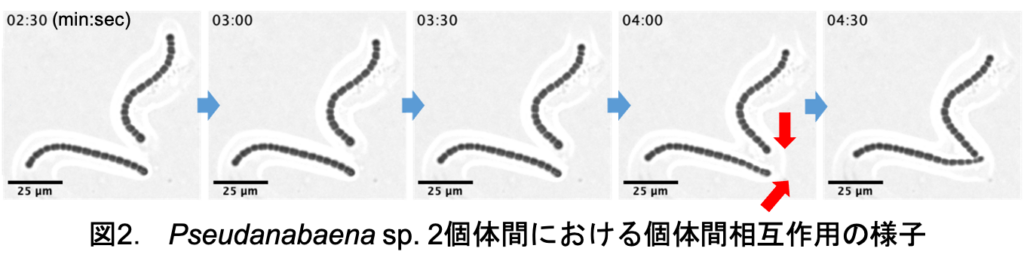
運動性シアノバクテリアにおける個体間相互作用の解析 杉田 篤俊
自然界において幾つかの生物種は群をなし集団生活を行う. 集団生活を行うことが生存競争に有利に働き, 種の繁栄に繋がると考えられている. 哺乳類や鳥類, 魚類に比べてより下等な生物であるバクテリアでも集団を形成することが知られている.
私の研究対象である運動性シアノバクテリアPseudanabaena sp. NIES-4403(以下Pseudanabaena sp. と表記)は粘液を分泌しながら滑走運動を行う. また, 特徴的な三種類の集団形態を形成し, 個体同士が相互に作用し合うことで集団形態を遷移する(図1). 本研究では集団が形成され始める瞬間に着目し集団形成のメカニズムを明らかにすることを目指す. Pseudanabaena sp. は個体同士の距離が一定以上近づくことで互いを引き込み合う相互作用を示す[1] . この相互作用が集団形成に関与すると仮定し, 個体間相互作用の実態を明らかにすべく運動解析を行う.
今回, Pseudanabaena sp. 2個体間における個体間相互作用の様子を位相差顕微鏡で撮影した(図2). 撮影画像について個体が他個体に引き付けられる際の運動速度や個体間距離, 方向を算出することで運動解析を行った.
その結果, 個体間距離に応じて引き付け方向に対する運動速度が上昇することが示された. また, 個体間相互作用によって運動速度が上昇し始める個体間距離を解析すると個体の周りを覆う粘液同士が融合した時点で個体間相互作用が生じ始める可能性が示唆された. 今後はPseudanabaena sp. の運動機構と個体間相互作用の関係を調査する予定である.
[1] 工藤亮太(2014)”運動性シアノバクテリアの個体間およびコロニー間における相互作用の解析”, 早稲田大学先進理工学部電気・情報生命工学科高松研究室, 2014年度修士論文
上皮細胞単体の運動と形態の基質依存性 岡本 直樹
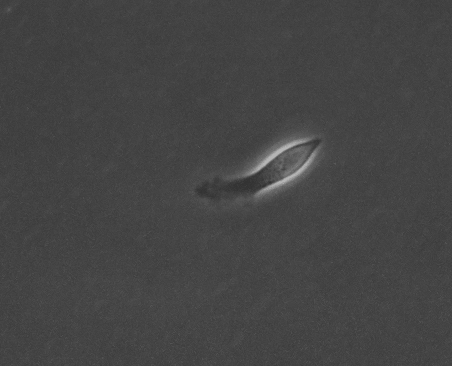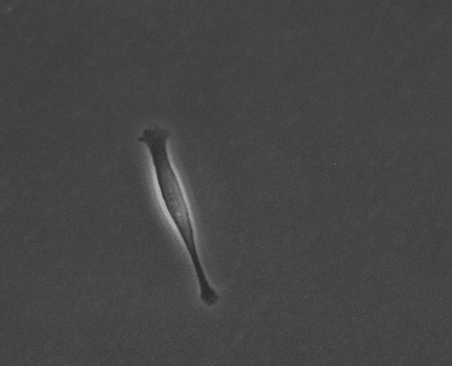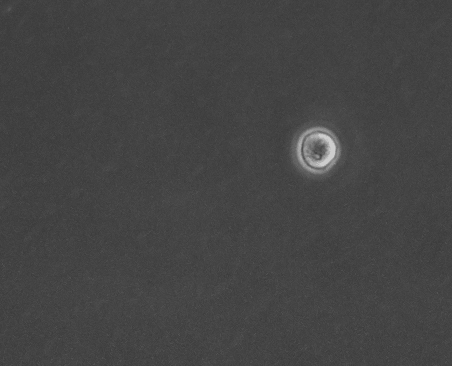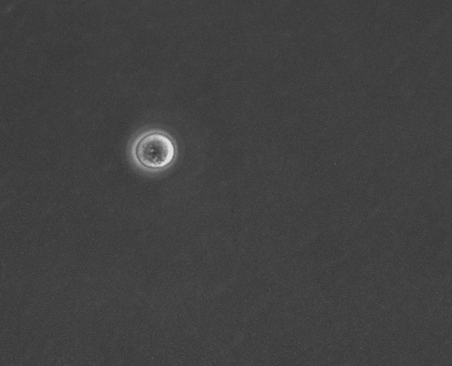
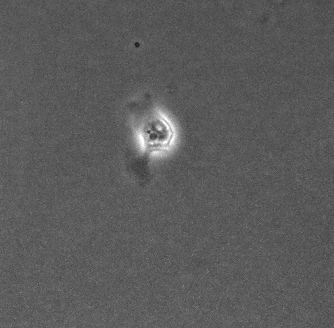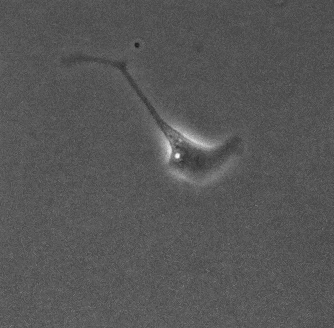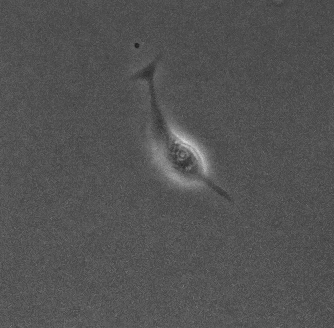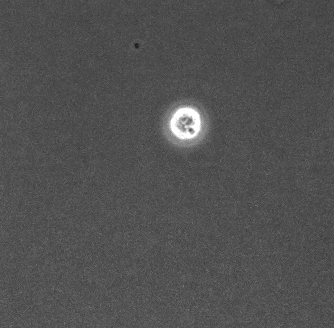
細胞外基質の環境は細胞の運動や形態の制御, さらには分化の誘導をするということが知られている. しかし, 本研究で扱うMDCK(Madin-Darby Canine Kidney)細胞は同一基質条件下でも孤立状態の細胞が多様な運動と形態を示す[1]. 孤立状態のMDCK細胞においても基質の環境が多様な形態を制御するのかを確かめ, そのメカニズムを明らかにすることを目的としている.
今回, コラーゲンを薄くコーティングした基質上とコーティングをしていない基質上でMDCK細胞を培養し, 実験を行った. 位相差顕微鏡で撮影した画像から形態分類, 面積などの形態指標の解析を行った.
その結果, コラーゲンをコーティングした細胞は過去に観察されていた形態(図1)に加え, 複雑で大きな形態が多く現れるようになった(図2). また, 直進性が高い形態も多く出現する傾向があることから運動性の上昇が示唆される. 今後は運動性についての解析や, 数理モデル(当研究室:八塚)と比較して細胞外基質の環境による細胞の運動と形態制御のメカニズムを調査していく予定である.
柴垣志文(2020),”孤立した上皮細胞の運動と形態の解析”早稲田大学大学院,先進理工学研究科,修士論文
誘引・忌避混合環境下における真正粘菌変形体の行動解析 入谷 涼
生物は様々な場面において最適な行動をとる. 変化する環境において, 周りを認識し適応させていくことは生きていくうえで重要である. 本研究では真正粘菌変形体 (以下, 粘菌と略称する) を用いて, 環境への適応について研究を行う.
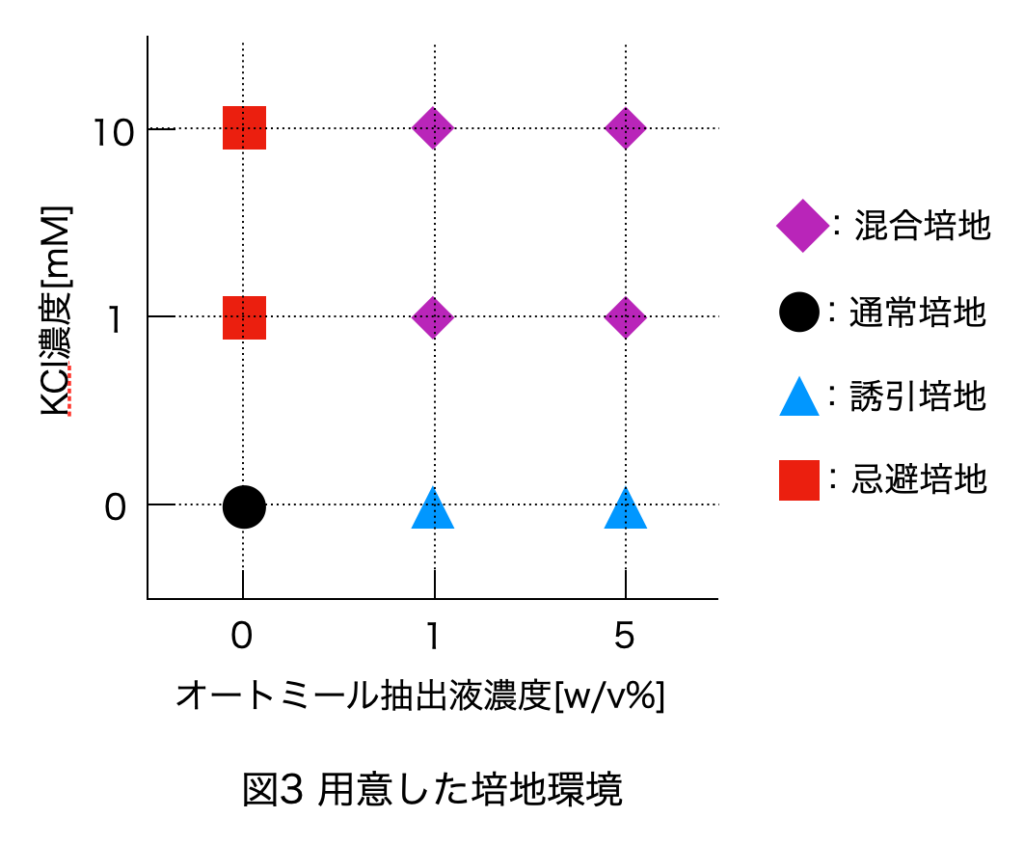
本研究の対象である粘菌は, 多核の単細胞生物であり輸送管ネットワークを構築する。粘菌は環境に応じて輸送管ネットワークの構造を変化させ、行動が変わるという特徴を持つ。先行研究から粘菌の好む環境 (誘引環境) では、メッシュ状のネットワーク構造を持ちゆっくりと進展する (図1) ことが、反対に粘菌が嫌う環境 (忌避環境) では、樹状のネットワーク構造を持ち素早く進展する (図2) ことがわかっている[1] 。本研究では、誘引や忌避の単独な環境ではなく混合した環境の方がより自然環境に近いのではないかと考え、混合環境での粘菌の行動を明らかにすることを目的としている。誘引環境と忌避環境を混合した培地 (図3) を用意して実験を行い、培地上での粘菌の様子を観察し、定量的に解析した。
実験の結果、粘菌が成長し面積を大きくしていく時、面積の変化率がそれぞれの環境で異なり、混合環境では誘引と忌避の二つの因子の影響を受けていることが示唆された。今後は粘菌の先端の進展速度や粘菌の特徴の一つでもある厚み振動に着目して、定量的解析をさらに進めていく所存である。
[1]A. Takamatsu, et al., (2009), Environment-dependent morphology in plas-modium of true slime mold Physarum polycephalumand network growth model, J. Theor. Biol., 256, 29-44.
周期的空間刺激による真正粘菌変形体の記憶 諸国 桜
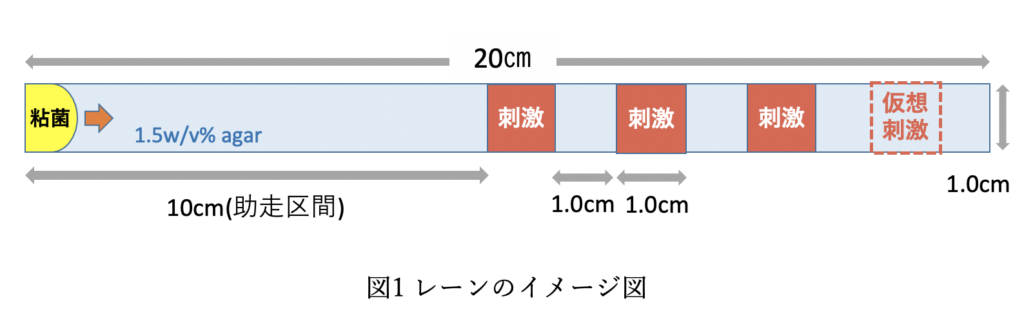
真正粘菌変形体(以下, 粘菌と略称する)は単細胞生物であるため脳や神経系を持たない。それにも関わらず, 粘菌は周期的な時間的刺激を記憶することがわかっている[1]. 粘菌が時間認知だけではなく空間認知も可能か検証することが本研究の目的である. そこで本研究では, まず空間的刺激の入力方法を確立するため, 刺激条件の選定を行った. 具体的には空間刺激を苦味物質であるキニーネとし刺激として機能する最適な濃度を探した. その後, 粘菌に対する周期的空間刺激入力の実験を行った.具体的には, 寒天培地で作成したレーンの後半に刺激物質を含んだ寒天ブロックを等間隔に埋め込み, そのレーン上を粘菌に渡らせ, 進行速度がどのように変化するか解析した.
実験の結果, 刺激下で進行速度が低下し仮想刺激下(刺激を与えていないが刺激が来るであろうと予測できる場所)で進行速度が低下した. これより周期的な空間刺激を粘菌は記憶する可能性が示唆された.
無刺激の通常レーンでも自発的な減速が見られるため, 仮想刺激下の減速が記憶によるものなのかを証明するために今後は統計的検証をしていく所存である.
[1]T. Saigusa, A.Tero, T. Nakagaki and Y. Kuramoto(2008), Amoebae Anticipate Periodic Events, Physical Review Letters, 100(1), 018101-1-4.
運動性シアノシアノバクテリアの個体とコロニーにおける運動の培地寒天濃度依存性解析 河本 優佑
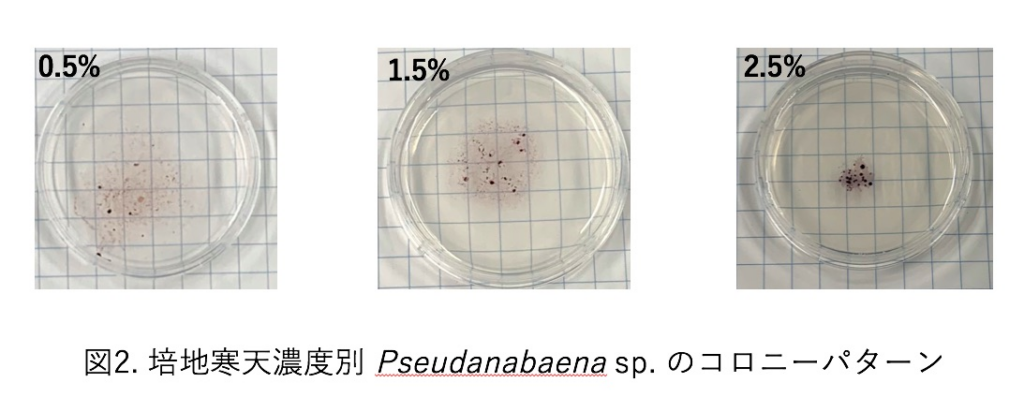
自然界において, 生物は周囲の環境に応じて振る舞いを変えることが知られている. これは魚類や哺乳類のみではなくサイズの小さいバクテリアでも見られる.
本研究で扱う運動性シアノバクテリアのPseudanabaena NIES-4403 (以下, Pseudanabaena sp.) は細胞が鎖状に連結して個体を形成しており, 粘液を分泌しながら滑走運動を行う. 各個体は他個体と相互作用することで三種類の集団形態をとる(図1). 環境に対する反応としてPseudanabaena sp.は培地の寒天濃度に応じてコロニーパターンが変化することが知られている(図2)[1]. 本研究ではこうした集団形態の形成、コロニーパターン変化のメカニズムおよびメリットを明らかにすることを目的とする.
寒天濃度は個体レベルの運動性にも影響を与えると仮定し実験を行なった. 寒天濃度の異なる三種類の培地を用意し、Pseudanabaena sp.の運動について位相差顕微鏡を用いて撮影することで速さを算出した.
現在までに培地の寒天濃度が高くなるにつれて個体および集団形態の運動速度が遅くなる傾向が得られ, 培地表面の水分が粘液と似たはたらきをしていることが示唆された. 今後は鎖状粒子モデルを用いて数値実験をすることで実験結果の再現を予定している[2].
[1] 深澤有貴(2012) “運動性シアノバクテリアのコロニーパターン形成のダイナミクス”, 早稲田大学大学院先進理工学研究科修士論文.
[2] 大友和理(2021) “運動性シアノバクテリアが形成する集団パターンの鎖状粒子モデルを用いた解析”, 早稲田大学大学院先進理工学研究科修士論文.
2023年度
運動性シアノバクテリアにおけるコロニーパターンの個体長依存性 黒川 理希
真正粘菌変形体における振る舞いのサイズ依存性 下津 怜士那
動きまわる細胞と組織の混合モデルの開発 福田 真大
2024年度
運動性シアノバクテリア集団形成の照射光波長依存性 松岡 早紀
生物は周囲の環境に応じて性質を変化させることで,過酷環境への耐性を高めている.生 存戦略は行動によるものや遺伝子によるものがあり,多様である.それぞれの環境に合わせた生存戦略が地球上の生命の多様性を支えているものと思われる.
本研究で扱う運動性シアノバクテリアPseudanabaena sp.NIES-4403(以下,Pseudanabaena sp.) は細胞を鎖状に連結させることで一つの個体を形成し,粘液分泌しながら滑走運動を行なっている. また,個体同士が相互作用することで,特徴的な三種類のコロニーパターンを形成する.(図1) Pseudanabaenaは原核光合成生物であり,周囲の光環境に応じて光受容体の構成色素を変化させる補色馴化という機能を有することが知られている[1].本研究では異なる波長で培養を行い,周囲の光環境による集団形成への影響を調べた.
本研究では,赤,青,緑の色補正フィルターでシャーレを覆い,コントロールと合わせて4種類の波長で培養を行なったところ,集団形成に違いがみられた.(図2)解析の結果,波長によって単個体の個体長や集団の速度,細胞数の増加率に影響があることが示唆された.それぞれの波長での集団の様子から,個体同士の相互作用にも影響を与えている可能性が考えられる.そこで 今後は,個体同士の相互作用を解析することで,Pseudanabaena の集団形成メカニズムをさらに詳しく調べる.
[1]成川礼(2013),“シアノバクテリアの光応答戦略-補色順化・走光性・細胞凝集をする光応答システム-“,植物科学最前線4;3-12
真正粘菌変形体の周期的空間記憶実験の数理モデル化 藪崎 広樹
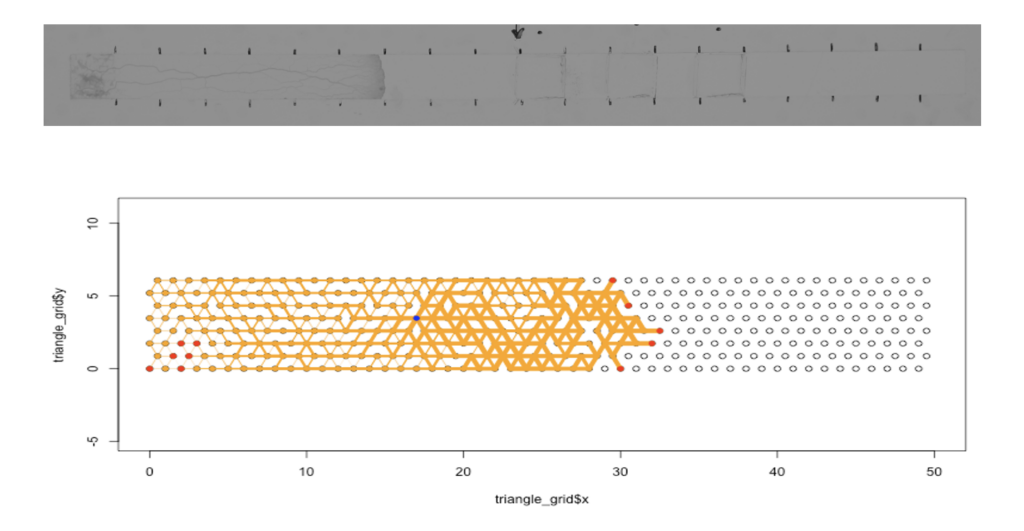
真正粘菌変形体(以下、粘菌と略称する)は多核の単細胞生物であり、約10𝜇𝑚~1𝑚の大きさを持つ。粘菌は細胞内部に輸送管ネットワークを形成し、誘引環境ではメッシュ状、忌避環境では樹状のネットワーク形態をとる。また、粘菌は単純な構造をもつ生物であるが、迷路を解くことができたり、環境の変化を「記憶」できるなど、知性を持つかのような振る舞いをすることが近年示唆されている。本研究では、粘菌の輸送管ネットワークに注目し、粘菌の記憶のメカニズムを明らかにすることを目的とする。そのために、粘菌が環境に応じてネットワークを形成する様子や周期的空間刺激を記憶する様子を再現できる数理モデルを構築する。
シアノバクテリアの個体長、速度、粘液幅の相互関係 小林 亮生
自然界でいくつかの生物種は生き残る生存戦略として集団生活を行い、群れを形成する。この集団行動は本研究の研究対象であるシアノバクテリアにも見られる。そこで、私の研究ではシアノバクテリアの個体長、移動速度、粘液の幅の相互関係について研究を行う。粘液とはシアノバクテリアが物質の表面を移動する運動、つまり滑走運動をする際に分泌するものである。粘液で囲まれた環境ではシアノバクテリアの移動速度が上昇していることが分かっている。個体長、移動速度、粘液幅の3つの相互関係を明らかにすることでシアノバクテリアの群れ形成メカニズムの調査に貢献する。
2025年度
大石 真帆
- 異なる硬さのコラーゲン培地における単一 MDCK 細胞の動態解析
小國 晴
- 寒天培地中必須塩濃度がシアノバクテリアの Disk 形態の運動性に及ぼす影響
須田 智晴
- 上⽪境界上の創傷治癒初期における細胞の振る舞い解析